| 1 | X-сцепленная адренолейкодистрофия |
| 2 | ААА синдром, Оллгрова синдром (ахалазия, алакримия, недостаточность надпочечников |
| 3 | Аарскога-Скотта cиндром |
| 4 | Абиотрофия сетчатки, тип Франческетти |
| 5 | Адреногенитальный синдром (врожденная гиперплазия коры надпочечников) |
| 6 | Азооспермия |
| 7 | Айкарди-Гутьереса синдром |
| 8 | Акродерматит энтеропатический |
| 9 | Аксенфельда-Ригера синдром |
| 10 | Алажиля синдром |
| 11 | Александера болезнь |
| 12 | Альбинизм глазокожный |
| 13 | Алькаптонурия |
| 14 | Альстрема синдром |
| 15 | Аменорея |
| 16 | Альфа-1-антитрипсина недостаточность |
| 17 | Ангельмана синдром |
| 18 | Андерсена синдром |
| 19 | Анемия Даймонда-Блекфена |
| 20 | Анеуплоидии |
| 21 | Аниридия |
| 22 | Антли-Бикслера синдром |
| 23 | Апера синдром |
| 24 | Арта cиндром |
| 25 | Артрогрипоз дистальный (синдром Фримена-Шелдона) |
| 26 | Атаксия Фридрейха |
| 27 | Атаксия, хорея, судороги и деменция |
| 28 | Атрофия зрительного нерва Лебера |
| Атрофия зрительного нерва с глухотой | |
| 30 | Аутоиммунный лимфопролиферативный синдром |
| 31 | Аутоиммунный полиграндулярный синдром I типа |
| 32 | Аутоимунный полиэндокринный синдром |
| 33 | Афазия первичная прогрессирующая |
| 34 | Ахондроплазия |
| 35 | Баллера-Герольда синдром |
| 36 | Банаян-Райли-Рувалькаба cиндром |
| 37 | Барде-Бидля (Ларенса-Муна) синдром |
| 38 | Барта cиндром |
| 39 | Барттера синдром |
| Бёрта-Хога-Дьюба синдром | |
| 41 | Бесплодие |
| 42 | Беста болезнь |
| 43 | Биотинидазы недостаточность |
| 44 | Блефарофимоз, обратный эпикант и птоз |
| 45 | Блоха-Сульцбергера синдром |
| 46 | Блума синдром |
| 47 | Боковой амиотрофический склероз |
| 48 | Боуэна-Конради синдром |
| 49 | Бранхио-окуло-фациальный синдром |
| 51 | Брахидактилия |
| 53 | Бьёрнстада синдром |
| 54 | Ваарденбурга синдром |
| 55 | Ваарденбурга-Шаха синдром |
| 56 | Ван дер Вуда синдром |
| 57 | Велокардиофациальный синдром |
| 58 | Вернера синдром |
| 59 | Видеманна-Беквита синдром, спорадическая нефробластома |
| 60 | Виллебранда болезнь |
| 61 | Вильсона-Коновалова болезнь |
| 62 | Вильямса cиндром |
| 63 | Вискотта-Олдрича cиндром |
| 64 | Вольмана болезнь, болезнь накопления эфиров холестерина |
| 65 | Вольфа-Хиршхорна синдром |
| 67 | Врожденная нечувствительность к боли с ангидрозом (врожденная сенсорная нейропатия с ангидрозом, HSAN4, CIPA) |
| 68 | Врожденной центральной гиповентиляции синдром |
| 69 | Вульгарный ихтиоз |
| 70 | Галактоземия тип I |
| 71 | Галактоземия тип II |
| 72 | Галактоземия тип III |
| 73 | Галактосиалидоз |
| 74 | Галлервордена-Шпатца болезнь |
| 75 | Ганглиозидоз GM1 тип 1,2,3 |
| 76 | Гастроинтестинальный полипоз |
| Гелеофизическая дисплазия | |
| 78 | Гемофилия |
| 79 | Гемохроматоз наследственный |
| 80 | Генитопателлярный синдром |
| 81 | Германски-Пудлака синдром |
| 82 | Герстманна-Штреусслера-Шейнкера болезнь |
| 83 | Гидроцефалия, обусловленная врожденнным стенозом Сильвиева водопровода |
| 84 | Гипер-IgD синдром |
| 85 | Гипер-IgM синдром |
| 86 | Гиперкалиемический периодический паралич |
| 87 | Гипероксалурия тип I |
| 88 | Гиперорнитинемии-гипераммониемии-гомоцитрулинурии синдром (ННН синдром) |
| 89 | Гипертрофическая кардиомиопатия |
| 90 | Гиперфенилаланинемия с дефицитом тетрагидробиоптерина |
| 91 | Гиперхолестеринемии |
| 92 | Гипогонадизм |
| 93 | Гипокалиемический периодический паралич |
| 94 | Гипоспадия |
| 95 | Гипотрихоз |
| 96 | Гипофосфатазия |
| 97 | Гипофосфатемический рахит |
| 98 | Гипохондроплазия |
| 99 | Гиппеля-Линдау синдром |
| 100 | Глазо-зубо-пальцевой синдром |
| 101 | Глаукома врожденная |
| 102 | Глаукома ювенильная открытоугольная |
| 103 | Гликогеноз 0 тип |
| 104 | Гликогеноз III типа |
| 105 | Гликогеноз IV типа |
| 106 | Гликогеноз IX типа |
| 107 | Гликогеноз Iа тип |
| 108 | Гликогеноз Iв тип |
| 109 | Гликогеноз V типа |
| 110 | Гликогеноз VI типа |
| 111 | Гликогеноз XI типа, Фанкони-Бикеля синдром |
| 112 | Гломеруоцитоз почек гипопластического типа |
| 113 | Глутаровая ацидурия тип 1 |
| 114 | Глутаровая ацидурия тип 2 |
| 116 | Гомоцистинурия |
| 117 | Гоше болезнь тип 1,2,3 |
| 118 | Грейга cиндром |
| 119 | Грисцелли cиндром |
| 120 | Дауна cиндром |
| 121 | Делеции хромосомы 1p36 синдром |
| 122 | Десмоидные опухоли |
| 123 | Дефицит гормона гипофиза, комбинированный |
| 124 | |
| 125 | Дефицит карнитина системный первичный |
| 126 | Дефицит фактора F12 |
| 127 | Джексона-Вейсса cиндром |
| 128 | Ди Джорджи cиндром |
| 129 | Диастрофическая дисплазия |
| 130 | Дисгенезия гонад |
| 131 | Дисплазия де ля Шапеля (Ателостеогенез) |
| 132 | Дисплазия Книста |
| 133 | Дистальная моторная нейропатия |
| 134 | Дистальная спинальная амиотрофия врожденная с параличом диафрагмы |
| 135 | Дисхондростеоз Лери-Вейлля |
| 136 | Дорфмана-Чанарина синдром |
| 137 | Жильбера cиндром |
| 138 | Жубер cиндром |
| 139 | Задержка полового созревания |
| 140 | Зандхоффа болезнь |
| 141 | Изовалериановая ацидемия |
| 142 | Инверсия пола |
| 143 | Ихтиоз буллезный |
| 144 | Ихтиоз врожденный аутосомно-рецессивный |
| 145 | Ихтиоз вульгарный |
| 146 | Ихтиоз, спастическая квадриплегия и умственная отсталость |
| 147 | Кампомелическая дисплазия |
| 148 | Канавана болезнь |
| 149 | Карбамолфосфатсинтетазы недостаточность |
| 150 | Карпентера cиндром |
| 151 | Кератита-ихтиоза-тугоухости cиндром |
| 152 | Кернса-Сейра синдром |
| 153 | Клайнфельтера cиндром |
| 154 | Клиппеля-Фейля cиндром |
| 155 | Коккейна cиндром |
| 156 | Комбинированный дефицит витамин K-зависимых факторов свертывания крови |
| 157 | Косолапость врожденная с или без дефицита длинных костей и/или зеркальной полидактилией |
| 158 | Костелло cиндром |
| 159 | Костная гетероплазия прогрессирующая |
| 160 | Коудена болезнь |
| 161 | Коффина-Лоури синдром |
| 162 | Кошачьего глаза синдром |
| 163 | Кошачьего крика синдром |
| 164 | Краббе болезнь |
| 165 | Краниометафизарная дисплазия |
| 166 | Краниосиностоз |
| 167 | Краниофациальной дисморфии-глухоты-ульнарной девиации кистей синдром |
| 168 | Крейтцфельда-Якоба болезнь |
| 169 | Криглера-Найара синдром |
| 170 | Крипторхизм |
| 171 | Крузона с черным акантозом синдром |
| 172 | Крузона синдром |
| 173 | Куррарино синдром |
| 174 | Ларинго-онихо-кутанный синдром |
| 175 | Лейкодистрофия с гипомиелинизацией |
| 176 | Лейкоэнцефалопатия с «исчезающим» белым веществом, детская атаксия с гипомиелинизацией |
| 177 | Лейкоэнцефалопатия с пораженим ствола мозга и высоким уровнем лактата при спектроскопии |
| 178 | Лейкоэнцефалопатия с субкортикальными кистами |
| 179 | Лейциноз (болезнь «с запахом кленового сиропа мочи» |
| 180 | Лермитт-Дуклос болезнь |
| 181 | Леша-Найяна синдром |
| 182 | Ли синдром |
| 183 | Ли-Фраумени синдром |
| 184 | Линча синдром (наследственный неполипозный рак толстой кишки) |
| 185 | Липодистрофия врожденная генерализованная |
| 186 | Липодистрофия семейная частичная |
| 187 | Липопротеин липазы недостаточность |
| 188 | Лоу синдром |
| 189 | Люджина — Фринса синдром |
| 190 | Макла-Уэллса синдром |
| 191 | Маклеода синдром |
| 192 | Малан синдром |
| 193 | Мандибулоакральная дисплазия с липодистрофией |
| 194 | Маннозидоз альфа |
| 195 | Маринеску-Шегрена синдром |
| 196 | Мартина-Белл, УО FRAXA Синдром |
| 197 | Маршалла-Смита синдром |
| 198 | Мевалоновая ацидурия |
| 200 | Метатропная дисплазия (OMIM 156530) |
| 201 | Метахроматическая лейкодистрофия |
| 202 | Метгемоглобинемия |
| 203 | Метилмалоновая ацидурия |
| 204 | Микрофтальм изолированный |
| 205 | Микрофтальм с катарактой |
| 206 | Микроцефалии с капиллярными мальформациями синдром |
| 207 | Миллера-Дикера синдром |
| 208 | Милроя болезнь (лимфедема наследственная) |
| 209 | Миоклоническая дистония |
| 210 | Миоклоническая эпилепсия Лабофа |
| 211 | Миопатия Броди |
| 212 | Миопатия Миоши |
| 213 | Миотоническая дистрофия |
| 214 | Миотония Томсена/Беккера |
| 215 | Митохондриальные гепатопатии |
| 216 | Митохондриальные заболевания, связанные с мутациями в гене POLG |
| 217 | Митохондриальные энцефаломиопатии, связанные с мутациями мтДНК |
| 218 | Митохондриальные энцефаломиопатии, связанные с мутациями ядерных генов |
| 219 | Множественная сульфатазная недостаточность |
| 220 | Множественной эндокринной неоплазии второго типа (МЭН2) cиндром |
| 221 | Множественные вывихи суставов, задержка роста, черепно-лицевые аномалии и врожденные пороки сердца |
| 222 | Множественных птеригиумов синдром |
| 223 | Множественных синостозов синдром |
| 224 | Молибденового кофактора недостаточность |
| 225 | Монилетрикс |
| 226 | Моуат-Вильсон cиндром |
| 227 | Муковисцидоз |
| 228 | Муколипидоз II, III типа |
| 229 | Мукополисахаридоз I типа |
| 230 | Мукополисахаридоз II типа |
| 231 | Мукополисахаридоз III А, В, С, D типа |
| 232 | Мукополисахаридоз IV A, B типа |
| 233 | Мукополисахаридоз VI типа |
| 234 | Мукополисахаридоз VII типа |
| 235 | Мышечная дистрофия врождённая, интегрин А7 негативная |
| 236 | Мышечная дистрофия врожденная, мерозин-негативная |
| 237 | Мышечная дистрофия врожденная, тип 1C |
| 238 | Мышечная дистрофия Дюшенна/Беккера |
| 239 | Мышечная дистрофия поясноконечностная |
| 240 | Мышечная дистрофия тип Фукуяма |
| 241 | Мышечная дистрофия Эмери-Дрейфуса |
| 242 | Мюнке синдром |
| 243 | Накопление нейтральных липидов с миопатией |
| 244 | Нарушение формирования пола |
| 245 | Нанизм MULIBRAY |
| 246 | Нарушения гликозилирования тип 1a, синдром Жакена |
| 247 | Нарушения гликозилирования тип Ib (ген MPI) |
| 248 | Наследственная моторно-сенсорная нейропатия (болезнь Шарко-Мари-Тута) тип I |
| 249 | Наследственная моторно-сенсорная нейропатия (болезнь Шарко-Мари-Тута) тип II |
| 250 | Наследственная нейропатия с подверженностью параличу от сдавления |
| 251 | Наследственная оптическая нейропатия Лебера |
| 252 | Наследственные глаукомы, аномалия Петерса, дермоид роговицы |
| 253 | Наследственный амилоидоз |
| 254 | Наследственный ангионевротический отек |
| 255 | Наследственный панкреатит |
| 256 | Невынашивание беременности |
| 257 | Наследственный рак желудка |
| 258 | Недостаточность N-ацетилглютаматсинтазы |
| 259 | Недостаточность длинноцепочечной 3-гидроксиацил-КоА-дегидрогеназы жирных кислот |
| 260 | Недостаточность короткоцепочечной ацил-КоА-дегидрогеназы жирных кислот |
| 261 | Недостаточность очень длинноцепочечной ацил-КоА дегидрогеназы жирных кислот |
| 262 | Недостаточность синтетазы голокарбоксилаз |
| 263 | Недостаточность среднецепочечной ацил-КоА-дегидрогеназы жирных кислот |
| 264 | Недостаточность сукцинил-КоА:3-кетоацил-КоА трансферазы |
| 265 | Незаращение родничков |
| 266 | Нейроаксональная дистрофия |
| 267 | Нейродегенерация с накоплением железа 4 |
| 268 | Нейромиотония и аксональная нейропатия |
| 269 | Нейрональный цероидный липофусциноз тип 1 |
| 270 | Нейрональный цероидный липофусциноз тип 2 |
| 271 | Нейросенсорная несиндромальная тугоухость |
| 272 | Нейрофиброматоз 1 и 2 типов |
| 273 | Нейтропения тяжёлая врождённая |
| 274 | Некетотическая гиперглицинемия |
| 275 | Некомпактного левого желудочка cиндром |
| 276 | Немалиновая миопатия |
| 277 | Нефронофтиз |
| 278 | Нефротический синдром |
| 279 | Ниймеген cиндром |
| 280 | Ниманна-Пика тип А и В болезнь |
| 281 | Ниманна-Пика тип С болезнь |
| 282 | Ногтей-надколенника синдром |
| 283 | Норри болезнь |
| 284 | Нунан синдром |
| 285 | Олигозооспермия тяжелой степени |
| 286 | Окулофарингеальная мышечная дистрофия |
| 287 | Опица GBBB синдром |
| 288 | Опица-Каведжиа синдром |
| 289 | Опухоль Вильмса |
| 290 | Орнитинтранскарбамилазы недостаточность |
| 291 | Ослера-Рендю-Вебера cиндром |
| 292 | Остеолиз карпотарзальный, мультицентрический |
| 293 | Остеопетроз рецессивный (мраморная болезнь костей) |
| 294 | Паллистера-Киллиана cиндром |
| 295 | Паллистера-Холла cиндром |
| 296 | Палочко-колбочковая дистрофия |
| 297 | Пантотенат киназы недостаточность |
| 298 | Парамиотония Эйленбурга |
| 299 | Патау cиндром |
| 300 | Пейтца-Егерса синдром |
| 301 | Пелицеуса-Мерцбахера болезнь |
| 302 | Пендреда Синдром |
| 303 | Первичная аутосомно-рецессивная микроцефалия, тип 5 |
| 304 | Первичная гипертрофическая остеоартропатия (пахидермопериостоз) |
| 305 | Первичная легочная гипертензия |
| 306 | Периодическая болезнь |
| 307 | Пигментная дегенерация сетчатки |
| 308 | Пикнодизостоз |
| 309 | Пирсона синдром |
| 310 | Пневмоторакс первичный спонтанный |
| 311 | Подколенного птеригиума cиндром |
| 312 | Полидактилия |
| 313 | Поликистоз почек |
| 314 | Помпе болезнь |
| 315 | Понтоцеребеллярная гипоплазия |
| 316 | Потоцки-Лупски cиндром |
| 317 | Почечная адисплазия |
| 318 | Прадера-Вилли Синдром |
| 319 | Преждевременная недостаточность яичников |
| 320 | Прогерия Хатчинсона-Гилфорда |
| 321 | Прогрессирующая наружная офтальмоплегия, АД и АР |
| 322 | Пропионовая ацидемия |
| 323 | Псевдоахондроплазия |
| 324 | Псевдоксантома эластическая |
| 325 | Пфайффера cиндром |
| 326 | Рабдомиолиз (миоглобинурия) |
| 327 | Рак молочной железы |
| 328 | Рак почки |
| 329 | Рак щитовидной железы. Синдром множественной эндокринной неоплазии второго типа (МЭН2) Синдром множественной эндокринной неоплазии второго типа (МЭН2) |
| 330 | Рак яичников |
| 331 | Ретинобластома |
| 332 | Ретиношизис |
| 333 | Ретта синдром |
| 334 | Рефсума болезнь |
| 335 | Ригидного позвоночника cиндром |
| 336 | Робинова синдром |
| 337 | Ротмунда-Томсена синдром |
| 338 | Рубинштейна-Тейби синдром |
| 339 | Семейная периодическая лихорадка |
| 340 | Семейный аденомоматозный полипоз, полипозный рак толстой кишки |
| 341 | Семейный внутрипеченочный холестаз 1 типа |
| 342 | Семейный внутрипеченочный холестаз 2 типа ( Баллера болезнь) |
| 343 | Семейный внутрипеченочный холестаз 3 типа |
| 344 | Семейный гемофагоцитарный лимфогистиоцитоз |
| 345 | Семейный медуллярный рак щитовидной железы |
| 346 | Семейный рак толстой кишки |
| 347 | Семейный холодовой аутовоспалительный синдром |
| 348 | Сениора-Локена синдром |
| 349 | Сенсорная полинейропатия (врожденная нечувствительность к боли) |
| 350 | Септо-оптическая дисплазия |
| 351 | Сетре-Чотзена синдром |
| 352 | Сиалидоз тип 1,2 |
| 353 | Сильвера-Рассела Синдром |
| 354 | Симпсона-Голаби-Бемель синдром |
| 355 | Синдром CADASIL, энцефалопатия с субкортикальными инфарктами |
| 357 | Синдром CINCA (холодовая лихорадка, синдром Мукле-Велса) |
| 358 | Синдром CRASH |
| 359 | Синдром ESC |
| 360 | Синдром LEOPARD |
| 361 | Синдром MASA |
| 362 | Синдром MNGIE |
| 363 | Синдром Ohdo, SBBYSS вариант |
| 364 | Синдром RAPADILINO |
| 365 | Синдром TAR |
| 366 | Синдром TRAPS (злокачественная гипертермия, амилоидоз почек) |
| 367 | Синдром тугоухости и атрофии зрительных нервов |
| 368 | Скапулоперонеальная миопатия |
| 370 | Смита-Лемли-Опитца синдром |
| 371 | Смит-Магенис синдром |
| 372 | Сотоса синдром |
| 373 | Спастическая параплегия Штрюмпеля |
| 374 | Спинальная амиотрофия типы I, II, III, IV |
| 375 | Спинальная и бульбарная амиотрофия Кеннеди |
| 376 | Спиноцеребеллярная атаксия |
| 377 | Спонгиоформная энцефалопатия с нейропсихическими проявлениями |
| 378 | Спондилокостальный дизостоз |
| 379 | Спондилоэпифизарная дисплазия (SEDT) |
| 380 | Стиклера синдром |
| 381 | Суперактивность фосфорибозилпирофосфат синтетазы |
| 382 | Талассемия beta |
| 383 | Тестикулярной феминизации синдром |
| 384 | Тея-Сакса болезнь |
| 385 | Тирозингидроксилазы недостаточность |
| 386 | Тирозинемия тип I |
| 387 | Торсионная дистония |
| 388 | Транспортера глюкозы недостаточность |
| 389 | Трихоринофалангеальный синдром |
| 390 | Тричера Коллинза-Франческетти синдром |
| 391 | Тромбоцитопения врожденная |
| 392 | Туберозный склероз |
| 393 | Умственная отсталость моногенная |
| 394 | Унферрихта-Лундборга болезнь |
| 395 | Уокера-Варбург синдром |
| 396 | Ушера синдром |
| 397 | Фабри болезнь |
| 398 | Фатальная семейная инсомния |
| 399 | Фацио-Лонде болезнь |
| 400 | Фелан-МакДермид синдром |
| 401 | Фенилкетонурия |
| 402 | Фибродисплазия оссифицирующая прогрессирующая |
| 403 | Фокальная кожная гипоплазия (Горлина-Гольца синдром) |
| 404 | Фокально-кортикальная дисплазия Тейлора |
| 405 | Фон Хиппель-Линдау Синдром |
| 406 | Фруктозо1,6 дифосфотазы недостаточность |
| 407 | Фукозидоз |
| 408 | Хайду-Чейни синдром |
| 409 | Хондродисплазия метафизарная тип Мак-Кьюсика |
| 410 | Хондродисплазия точечная Конради-Хюнермана |
| 411 | Хондрокальциноз |
| 412 | Хореоатетоз, гипотиреоидизм и неонатальная дыхательная недостаточность |
| 413 | Хорея Гентингтона |
| 414 | Хорея доброкачественная наследственная |
| 415 | Хороидермия |
| 416 | Хромосомные болезни |
| 417 | Хроническая гранулематозная болезнь |
| 418 | Х-сцепленная агаммаглобулинемия |
| 419 | Х-сцепленный лимфопролиферативный синдром (болезнь Дункана, синдром Пуртильо) |
| 420 | Х-сцепленный моторный нистагм |
| 421 | Х-сцепленный тяжелый комбинированный иммунодефицит |
| 422 | Целвегера синдром |
| 423 | Центронуклеарная миопатия |
| 424 | Цереброокулофациоскелетный синдром |
| 425 | Цистиноз |
| 426 | Цистиноз нефропатический |
| 427 | Цитруллинемия тип 1 |
| 428 | Шварца-Джампела синдром |
| 429 | Швахмана-Даймонда синдром |
| 430 | Шегрена-Ларссона синдром |
| 431 | Шерешевского-Тернера синдром |
| 432 | Широкого водопровода преддверия синдром |
| 433 | Шпринтцена-Гольдберга синдром |
| 434 | Штаргардта болезнь |
| 435 | Эдвардса синдром |
| 436 | Экзостозы множественные |
| 437 | Эксудитивная витреохореорстинальная дистрофия |
| 438 | Эктодермальная ангидротическая дисплазия |
| 439 | Эктодермальная гидротическая дисплазия |
| 440 | Эктопия хрусталика |
| 442 | Эллерса-Данло синдром |
| 443 | Эпилепсия прогрессирующая миоклоническая |
| 444 | Эпифизарная дисплазия, множественная |
| 445 | Эритрокератодермия |
| 446 | Эритроцитоз рецессивный |
| 447 | Эскобара cиндром |
Медицинская генетика
Аннотация Ожирение является важнейшим фактором риска развития сахарного диабета 2 типа (СД2).![]() Гепатоцитарный ядерный фактор 1 β (HNF1B) контролирует глюкостатическую функцию островков Лангерганса поджелудочной железы и ассоциирован с развитием СД2 в европейской и азиатской популяциях. Однако исследований, оценивающих роль генетических вариантов HNF1B в формировании предрасположенности к СД2 в русской популяции, на сегодняшний день не проводилось. Целью настоящей работы стало изучение ассоциации полиморфного варианта rs4430796 (A>G) в интроне гена HNF1B с показателями гликемического профиля и редокс-гомеостаза, а также риском развития СД2 у жителей Центральной России, с учетом их пола и индекса массы тела. В исследование включено 3206 человек, из них 1579 больных СД2 и 1627 условно здоровых добровольцев. Генотипирование проводили с использованиеми технологии iPLEX на геномном времяпролетном масс-спектрометре MassArray 4 (Agena Bioscience). Впервые в русской популяции установлена взаимосвязь полиморфизма rs4430796 гена HNF1B с повышенным риском развития СД2 (OR 1,24, 95CI 1,05-1,47, р=0,011).
Гепатоцитарный ядерный фактор 1 β (HNF1B) контролирует глюкостатическую функцию островков Лангерганса поджелудочной железы и ассоциирован с развитием СД2 в европейской и азиатской популяциях. Однако исследований, оценивающих роль генетических вариантов HNF1B в формировании предрасположенности к СД2 в русской популяции, на сегодняшний день не проводилось. Целью настоящей работы стало изучение ассоциации полиморфного варианта rs4430796 (A>G) в интроне гена HNF1B с показателями гликемического профиля и редокс-гомеостаза, а также риском развития СД2 у жителей Центральной России, с учетом их пола и индекса массы тела. В исследование включено 3206 человек, из них 1579 больных СД2 и 1627 условно здоровых добровольцев. Генотипирование проводили с использованиеми технологии iPLEX на геномном времяпролетном масс-спектрометре MassArray 4 (Agena Bioscience). Впервые в русской популяции установлена взаимосвязь полиморфизма rs4430796 гена HNF1B с повышенным риском развития СД2 (OR 1,24, 95CI 1,05-1,47, р=0,011). Стратифицированный анализ по полу обнаружил, что выявленная ассоциация характерна только для женщин с избыточной массой тела (OR 1,54, 95CI 1,06-2,22, р=0,02) и ожирением (OR 2,07, 95CI 1,14-3,77, р=0,047) и отсутствует у лиц с нормальной массой тела вне зависимости от пола. Изучаемый SNP ассоциирован с повышенным содержанием перекиси водорода (р=0,012) и более низким уровнем общего глутатиона плазмы (р=0,041) у женщин, тогда как у мужчин с СД2 генотип G/G связан со снижением концентрации С-пептида (р=0,004) и повышением концентрации глюкозы крови (р=0,015). Биоинформатический анализ подтвердил отрицательный эффект аллеля G на экспрессию HNF1B, а также выявил его связь с гиперметилированием гена в различные периоды жизни, что обусловливает низкую экспрессию гена HNF1B у носителей минорного аллеля rs4430796-G. Таким образом, нами впервые установлено, что полиморфный вариант гена HNF1B rs4430796 ассоциирован с предрасположенностью к СД2 в русской популяции, при этом его связь с заболеванием имеет пол-специфический характер и зависит от индекса массы тела.
Стратифицированный анализ по полу обнаружил, что выявленная ассоциация характерна только для женщин с избыточной массой тела (OR 1,54, 95CI 1,06-2,22, р=0,02) и ожирением (OR 2,07, 95CI 1,14-3,77, р=0,047) и отсутствует у лиц с нормальной массой тела вне зависимости от пола. Изучаемый SNP ассоциирован с повышенным содержанием перекиси водорода (р=0,012) и более низким уровнем общего глутатиона плазмы (р=0,041) у женщин, тогда как у мужчин с СД2 генотип G/G связан со снижением концентрации С-пептида (р=0,004) и повышением концентрации глюкозы крови (р=0,015). Биоинформатический анализ подтвердил отрицательный эффект аллеля G на экспрессию HNF1B, а также выявил его связь с гиперметилированием гена в различные периоды жизни, что обусловливает низкую экспрессию гена HNF1B у носителей минорного аллеля rs4430796-G. Таким образом, нами впервые установлено, что полиморфный вариант гена HNF1B rs4430796 ассоциирован с предрасположенностью к СД2 в русской популяции, при этом его связь с заболеванием имеет пол-специфический характер и зависит от индекса массы тела.
ФГБНУ «НИИ АГиР им.Д.О.Отта» :: ФГБНУ «НИИ АГиР им.Д.О.Отта»
Сведения об образовании
ВУЗ: Львовский государственный медицинский институт, 1963 г. Специальность по диплому. По специальности лечебного факультета. Квалификация: врач.
Аспирантура: НИИ экспериментальной медицины АМН СССР, г. Ленинград (1963-1966 гг.).
Защита кандидатской диссертации в 1966 г. (НИИЭМ АМН СССР, г. Ленинград).
Защита докторской диссертации в 1976 г. (НИИЭМ АМН СССР, г. Ленинград).
Курсы повышения квалификации, сертификационные курсы
По специальности Генетика, 1996, 2001, 2006, 2011 гг. на кафедре медицинской генетики СПб МАПО.
Высшая квалификационная категория по специальности «Генетика» с 1998 г.
Сведения о наградах, премиях, почетных званиях
Диплом и премия им. акад. А.А.Баева «За разработку научных основ и внедрение в клиническую практику методов генодиагностики наиболее распространенных социально значимых наследственных болезней», 1995 г.
Нагрудный знак «200 лет Научно-исследовательскому институту акушерства и гинекологии им.Д.О.Отта РАМН», 1997 г.
Почетная Грамота Российской Академии Медицинских Наук «За плодотворный труд по развитию медицинской науки и здравоохранения», 1997 г.
Диплом Координационного Межведомственного Совета по Приоритетному направлению Науки о Жизни и биотехнология, 1998 г.
Золотая медаль 1-го Международного салона инноваций и инвестиций, 1991 г.
Грамота Военно-медицинской академии «За добросовестное выполнение обязанностей и проявленную активность в учебной, методической и воспитательной работе по итогам 2003/2004 учебного года», 2004 г.
Почетная грамота Министерства Здравоохранения и Социального Развития РФ, 2005 г.
Диплом и памятная медаль С.Н.Давиденкова Российского общества медицинских генетиков, 2005 г.
Нагрудный знак «Отличник здравоохранения» МЗСР РФ, 2007 г.
Премия имени И.П. Павлова Правительства Санкт-Петербурга и Санкт-Петербургского научного центра РАН в области науки и медицины, 2008 г.
Диплом премии РАМН имени С.Н.Давиденкова за лучшую работу по медицинской генетике, 2009 г.
Грамота РАМН «За плодотворный труд по развитию медицинской науки и здравоохранения», 2010 г.
Грамота Губернатора Санкт-Петербурга, 2010 г.
Почетная грамота Комитета по здравоохранению Правительства Санкт-Петербурга, 2010 г.
Благодарность Законодательного собрания Санкт-Петербурга, 2010 г.
Почетный Диплом Европейской научно-промышленной палаты «За фундаментальный вклад в биологию и медицину», 2012 г.
Диплом Российского общества медицинских генетиков, 2014 г.
Юбилейная медаль «50 лет Вавиловского общества генетиков и селекционеров», 2017 г.
Медаль «За выдающийся вклад в развитие медицинской генетики», учрежденная ФГБНУ «Медико-генетический научный центр», 2018 г.
Участие в грантах (текущие проекты на 2018 г.)
Грант РФФИ 16-04-01438 (2016-2018). Специфичность паттернов 5-гидроксиметилцитозина в геноме клеток эмбриональных и экстраэмбриональных тканей человека при нормальном развитии и эмбриональных потерях.
Грант РНФ №14-15-00737 (2014-2018). «Исследование молекулярно-генетических, эпигенетических и цитогенетических особенностей функции генома клеток миометрия и эндометрия матки в норме и патологии».
Руководство/консультации кандидатскими и докторскими диссертациями
Кандидатские диссертации
Джолахава Спиридон Спиридонович. Оценка повреждающего действия некоторых лекарственных препаратов на эмбриональное развитие в эксперименте. (Тбилиси, НИИАГ, 1987).
Ветух Виктор Александрович. Изучение роли опосредованного действия радионуклидов различной тропности и генетическом повреждении половых клеток. (Ленинград, ЦНИРИ МЗ СССР, 1991).
Нефедова Татьяна Николаевна. Развитие методов трансплантации эмбрионов млекопитающих. (Москва,Ин-т истории естествознания и техники АН СССР, 1992).
Лившиц Людмила Аврамовна. Полиморфизм ДНК области q31 хромосомы 7 у человека в семьях высокого риска муковисцидоза и региональных популяциях Украины. (Киев, Институт молекулярной биологии и генетики АН Украины, 1992).
Иващенко Татьяна Эдуардовна. Идентификация мутаций и ПДРФ анализ ДНК локусо, сцепленных с геном муковисцидоза, в некоторых популяциях, семьях высокого риска и у больных. (Москва, МГНЦ РАМН, 1992).
Лебедев Владимир Михайлович. Сравнительный анализ методов получения плодного материала в первом триместре беременности для пренатальной диагностики наследственных болезней. (СПб, НИИАГ им.Д.О.Отта РАМН, 1993).
Бобоев Кодиржон Тухтабоевич. Молекулярно-генетический анализ гемофилии А в Узбекистане. (Ташкент, Онкологический научный центр САО АМН, 1993).
Агбангла Клемент. Анализ полиморфизма и идентификация мутаций гена трансмембранного регуляторного белка муковисцидоза в популяциях человека. (СПб, СПбГУ, 1993).
Гимбовская Светлана Дмитриевна. Рестрикционный полиморфизм ДНК в области «гена муковисцидоза», ассоциация его вариантов с заболеванием и мутациями гена в республике Молдова. (Москва, МГНЦ РАМН, 1993).
Ракишева Зере Баяновна. Диагностические основы для медико-генетической профилактики синдрома Мартина-Белла (С. Fra(Xq27). (Москва, МГНЦ РАМН, 1993).
Fra(Xq27). (Москва, МГНЦ РАМН, 1993).
Бакунин Сергей Викторович. Акушерские аспекты биопсии плаценты в пренатальной диагностике хромосомных и генных болезней. (СПб, НИИАГ им.Д.О.Отта РАМН, 1994).
Живац Зинаида Михайловна. Клинико-генетическая характеристика фенилкетонурии у детей у детей у детей в Одесской области. (Одесса, Медицинский институт им. Н.И.Пирогова, 1994).
Кащеева Татьяна Константиновна. Некоторые белки амниотической жидкости. (СПб, ИЭМ РАМН, 1995).
Горностаева Наталья Юрьевна. Профилактика гемофилии А у детей с использованием клинических и молекулярно-генетических методов. (Одесса, Медицинский институт им. Н.И.Пирогова, 1996).
Асеев Михаил Владимирович. Молекулярно-генетический анализ генов факторов VIII и IX в некоторых популяциях и семьях с гемофилией А и В. (СПб, Росссийский НИИ гематологии и трансфузиологии МЗ МП РФ, 1996).
Ковалева Людмила Федоровна. Особенности клинического течения муковисцидоза у детей гетро- и гомозигот по delF508 по данным Центра муковисцидоза г. Санкт-Петербурга. (СПб, Государственный научный центр пульмонологии, 1997).
Санкт-Петербурга. (СПб, Государственный научный центр пульмонологии, 1997).
Нагорная Ирена Игоревна. Клинический, молекулярно-генетический и цитогенетический анализ у пациентов с нарушениями половой дифференцировки. (СПб, Государственная педиатрическая медицинская академия, 1998).
Бакай Марина Анатольевна. Молекулярно-генетический анализ гена трансмембранного регуляторного белка муковисцидоза у больных Санкт-Петербурга. Корреляция генотип-фенотип. (СПб, ИЭМ РАМН, 1998).
Скакун Владимир Николаевич. Сравнительный анализ полиморфизма некоторых коротких тандемных повторов в трех популяциях человека и их использование для генетической идентификации личности. (СПб, СПбГУ, 1999).
Шауи Абдельрхани. Анализ частот мутантных аллелей трех генов предрасположенности к мультифакториальным заболеваниям в популяции и у больных Северо-Западного региона России. (СПб, СПбГУ, 1999).
Карева Инна Валентиновна. Факторы генетического и акушерского риска в пренатальной диагностике хромосомных болезней. (СПб, НИИАГ им.Д.О.Отта РАМН, 1999).
(СПб, НИИАГ им.Д.О.Отта РАМН, 1999).
Глазков Павел Борисович. Изучение некоторых особенностей экспрессии гена CFTR и разработка экспериментальных подходов к генотерапии муковисцидоза (СПб, СПбГУ, 2000).
Ледащева Татьяна Алексеевна. Псевдогипертрофическая мышечная дистрофия Дюшенна/Беккера (СПб, МАПО, 2000).
Малышева Ольга Викторовна. Молекулярно-генетический анализ генов IT-15 и DPMK в популяциях и в семьях с миотонической дистрофией и хореей Гентингтона. (СПб, СПбГУ, 2000).
Баранов Александр Николаевич. Исследования невирусных способов доставки генных кострукций для генотерапии миодистрофии Дюшенна (СПб, СПбГУ, 2000).
Беспалова Олеся Николаевна. Оценка роли генетических факторов в привычном невынашивании беременности ранних сроков. (СПб, НИИАГ им.Д.О.Отта РАМН, 2001).
Сидилева Ольга Геннадьевна. Полиморфные аллели генов, ассоциированных с патогенезом атопической формы бронхиальной астмы у жителей Северо-Запада России (СПб, СПбГУ, 2002).
Гагарина Александра Владимировна. Плацентарная недостаточность при повышенном содержании альфа-фетопротеина и хорионического гонадотропина в крови беременных женщин (СПб, НИИАГ им.Д.О.Отта РАМН, 2004).
Плацентарная недостаточность при повышенном содержании альфа-фетопротеина и хорионического гонадотропина в крови беременных женщин (СПб, НИИАГ им.Д.О.Отта РАМН, 2004).
Киселев Антон Вячеславович. Невирусные носители доставки ДНК в клетки млекопитающих с целью генотерапии миодистрофии Дюшенна (СПб, СПбГУ, 2005).
Некрасова Екатерина Сергеевна. Комбинированный ультразвуковой и биохимический скрининг хромосомной патологии плода в первом триместре беременности у беременных женщин Северо- Западного региона России (СПб, НИИАГ им.Д.О.Отта РАМН, 2005).
Лязина Лидия Викторовна. Клинические и цитогенетические исследования у детей и подростков с аномалиями половых хромосом (СПб, ГОУВПО «Санкт-Петербургская государственная педиатрическая медицинская академия» Росздрава, 2005).
Глотов Олег Сергеевич. Анализ полиморфизма генов сердечно-сосудистой системы и системы детоксикации в различных возрастных группах Санкт-Петербурга (СПб, СПбГУ, 2007).
Егорова Анна Алексеевна. Линейные и разветвленные поли- и олигопептиды на основе лизина как стредства доставки генных конструкций в клетки млекопитающих (СПб, СПбГУ, 2009).
Бикмуллина Дина Рустемовна. Молекулярно-генетические и биохимические показатели риска коагулопатического кровотечения у беременных с гестозом и его профилактика (СПб, НИИАГ им. Д.О. Отта СЗО РАМН, 2010).
Махрова Ирина Александровна. Наследственная предрасположенность к метаболическому синдрому у детей (СПб, ГБОУ ВПО СПбГПМА, 2011).
Москаленко Михаил Викторович. Полиморфизм ряда генов метаболизма костной ткани и остеопороз у человека (СПб, СПбГУ, 2011).
Железнякова Галина Юрьевна Влияние характера метилирования геномной ДНК и числа копий гена SMN2 на тяжесть спинальной мышечной атрофии (СПб, СПбГУ, 2014).
Шильникова Евгения Михайловна. Генетические и эпигенетические особенности генома сперматозоида и их влияние на раннее эмбриональное развитие человека (СПб, СПбГУ, 2015).
Вашукова Елена Сергеевна. Молекулярно-генетические аспекты развития гестоза у женщин Северо-Западного региона России (СПб, СПбГУ, 2017).
Докторские диссертации
Павлов Геннадий Валерьевич. Муковисцидоз, его эволютивные формы и различные периоды детства у взрослых (Екатеринбург, Уральский Государственный Медицинский Институт, 1993).
Муковисцидоз, его эволютивные формы и различные периоды детства у взрослых (Екатеринбург, Уральский Государственный Медицинский Институт, 1993).
Горбунова Виктория Николаевна. Биохимические и молекулярные аспекты пренатальной диагностики некоторых врожденных и наследственных заболеваний (дефекты заращения нервной трубки, муковисцидоз, миодистрофия Дюшенна, гемофилия А). (Москва, МГНЦ РАМН, 1996).
Михайлов Антон Валерьевич. Клинико-патофизиологические аспекты внутриматочных вмешательств в целях диагностики и лечения врожденных и наследственных заболеваний плода. (СПб, НИИАГ им.Д.О.Отта РАМН, 1999).
Иващенко Татьяна Эдуардовна. Муковисцидоз: молекулярный анализ гена, разработка новых подходов диагностики и генотерапии. (Москва, МГНЦ РАМН, 2000).
Кузнецова Татьяна Владимировна. Комплексный подход к цитогенетике эмбрионального развития человека. (СПб, СПбГУ, 2000).
Кащеева Татьяна Константиновна. Пренатальный биохимический скрининг – система, принципы, клинико-диагностические критерии, алгоритмы. (СПб, Всерос. центр экстрен. и радиац. медицины им. А.М. Никифорова МЧС России, 2009).
(СПб, Всерос. центр экстрен. и радиац. медицины им. А.М. Никифорова МЧС России, 2009).
Беспалова Олеся Николаевна. Генетические факторы риска невынашивания беременности (СПб, НИИАГ им. Д.О. Отта СЗО РАМН, 2009).
Глотов Андрей Сергеевич Генетические и средовые факторы риска развития гестоза у женщин с артериальной гипертензией и метаболического синдрома у детей (СПбГУ, 2017).
Оппонирование кандидатских/докторских диссертаций
Кандидатские диссертации
Архангельская Ирина Борисовна. Чувствительность in vitro доимплантационных зародышей мыши к повреждающему действию некоторых эмбриотоксических факторов — НИИ экспериментальной медицины, Санкт-Петербург, 1996.
Платонова Наталья Андреевна. Совместное участие рецептора церулоплазмина и медьтранспортной АТРазы, продукта гена болезни Менкеса, в связывании церулоплазмина – НИИЭМ РАМН Санкт-Петербург, 2000.
Якупова Эльвира Венеровна. Молекулярно-генетические механизмы формирования гемобластозов у детей — Башкир. гос. мед. ун-т. — Уфа, 2004.
гос. мед. ун-т. — Уфа, 2004.
Игнатович Ирина Анатольевна. Катионные пептиды как средство переноса ДНК в клетки млекопитающих / С.-Петерб. гос. ун-т. — Санкт-Петербург, 2005.
Мудрак Ольга Станиславовна. Архитектура хромосом в ядре сперматозоида человека / Ин-т цитологии РАН. — Санкт-Петербург, 2006.
Капустин Сергей Игоревич. Молекулярно-генетические аспекты патогенеза венозного тромбоэмболизма [Место защиты: Рос. науч.-исслед. ин-т гематологии и трансфузиологии РФ]. — Санкт-Петербург, 2007.
Соколова Анастасия Владимировна. Дифференцировка и регенерация скелетных мышц мышей mdx после клеточной терапии стволовыми клетками костного мозга [Место защиты: Ин-т цитологии РАН]. — Санкт-Петербург, 2010.
Плюснина Екатерина Николаевна. Влияние сверхэкспрессии гена D-GADD45 в нервной системе Drosophila melanogaster на продолжительность жизни, возрастзависимые физиологические показатели и стрессоустойчивость [Место защиты: С.-Петерб. ин-т биорегуляции и геронтологии Сев.-Зап. отд-ние РАМН]. — Санкт-Петербург, 2011.
— Санкт-Петербург, 2011.
Васькова Евгения Андреевна. Модификации хроматина при инактивации X-хромосомы у грызунов [Место защиты: Ин-т цитологии и генетики СО РАН]. — Новосибирск, 2014.
Титов Борис Васильевич. Полиморфизм генов, белковые продукты которых играют роль в развитии воспаления, при ишемическом инсульте [Место защиты: Научно-исследовательский институт экспериментальной медицины ФГБУ СЗО РАМН].- Санкт-Петербург, 2014.
Домнина Алиса Павловна. Эндометриальные стволовые клетки: получение, характеристика и применение для стимуляции развития эндометрия крыс [Место защиты: Ин-т цитологии РАН, СПб]. — Санкт-Петербург, 2014.
Докторские диссертации
Яковлев Александр Федорович. Структурно-функциональные особенности генома клеток сельскохозяйственных животных – ВНИИРГЖ, Ленинград, 1979.
Кириллова Ирина Алексеевна. Патологическая анатомия и патогенез пороков развития в пренатальном онтогенезе человека — Мед. академия последипломного образования — Санкт-Петербург, 1994.
Паткин Евгений Львович. Динамика структурно-функциональной организации метафазных и интерфазных хромосом в раннем эмбриогенезе млекопитающих в исследовании in situ — НИИ экспериментальной медицины. — Санкт-Петербург, 1995.
Залетаев Дмитрий Владимирович. Цитогенетический и молекулярно-генетический анализ критических районов хромосом, повреждаемых при менделирующих синдромах МВПР — Медико-генетический науч. центр. — Москва, 1996.
Мандельштам Михаил Юрьевич. Молекулярно-генетический анализ моногенных форм атеросклероза и рака молочной железы у жителей Санкт-Петербурга / Науч.-исслед. ин-т эксперим. медицины РАМН. — Санкт-Петербург, 2005.
Даев Евгений Владиславович. Генетические последствия ольфакторных стрессов у мышей / С.-Петерб. гос. ун-т. — Санкт-Петербург, 2006.
Руденко Дмитрий Игоревич. Лице-лопаточно-плечевая мышечная дистрофия и ее связь с лице-лопаточно-перонеальной мышечной дистрофией : клиника и генетика [Место защиты: С.-Петерб. гос. мед. ун-т им. И.П. Павлова]. — Санкт-Петербург, 2009.
— Санкт-Петербург, 2009.
Сироткина Ольга Васильевна. Молекулярно-генетические механизмы активации тромбоцитов и чувствительности к антиагрегантным препаратам [Место защиты: Всерос. центр экстрен. и радиац. медицины им. А.М. Никифорова МЧС России]. — Санкт-Петербург, 2011.
Пчелина Софья Николаевна. Молекулярно-генетические основы наследственных форм болезни Паркинсона [Место защиты: Санкт-Петербургский государственный университет]. — Санкт-Петербург, 2012.
Саранцева Светлана Владимировна. Роль генов пресенилина 1 и белка предшественника амилоида в дисфункции синапсов при болезни Альцгеймера. [Место защиты: Санкт-Петербургский государственный университет]. — Санкт-Петербург, 2012.
Боринская Светлана Александровна. Генетическая адаптация популяций человека к природным и антропогенным факторам среды [Место защиты: Ин-т общ. генетики им. Н.И. Вавилова РАН]. — Москва, 2013.
Черных Вячеслав Борисович. Аномалии половых хромосом при нарушениях формирования пола и репродукции человека [Место защиты: Мед. -генет. науч. центр РАМН]. — Москва, 2015.
-генет. науч. центр РАМН]. — Москва, 2015.
Вохмянина Наталья Васильевна. Лабораторная диагностика целиакии : принципы и алгоритмы [Место защиты: Всерос. центр экстрен. и радиац. медицины им. А.М. Никифорова МЧС России]. — Санкт-Петербург, 2016.
Внешнее совмещение
Санкт-Петербургский государственный университет, кафедра генетики и биотехнологии, профессор (0,5).
Военно-медицинская академия, кафедра детских болезней, профессор (0,5).
Кафедры ИДПО
Контактная информация:
Адрес: г. Уфа, ул. Гафури, 74, Республиканский медико-генетический центр,2 этаж.
Почтовый адрес e-mail: [email protected]
Положение о кафедре
История кафедры
Многолетние и разносторонние исследования наследственных нервно-мышечных заболеваний, проводимые на кафедре ассистентом Маргариты Евгеньевны Третьяковой, создали предпосылки для открытия в 1970 г. под научно-методическим руководством кафедры нервных болезней БГМИ медико-генетической консультации (МГК) при Республиканской клинической больнице им. Г.Г.Куватова (РКБ) и начала преподавания медицинской генетики.
Третьякова М.Е., невролог, кандидат медицинских наук (1962 г.), окончила с отличием БГМИ.
В 1946-1949 гг. – зав. Районным отделом здравоохранения Благоварского района и терапевт ЦРБ. В1949-1952 гг. инспектор и начальник группы здравоохранения Совета Министров БАССР, одновременно – клинический ординатор кафедры нервных болезней БГМИ. В 1952-1954 гг. – ординатор, а в 1954-1962 гг. заведующая неврологическим отделением РКБ. В 1962-1964 гг. – научный сотрудник УфНИИ гигиены и профзаболеваний. С 1964 по 1996 гг. – ассистент кафедры неврологии БГМИ. Изучала заболеваемость клещевым энцефалитом в Башкирии, заболеваемость рабочих нефтеперерабатывающих и других заводов; наследственные заболевания нервно-мышечной системы. Имеет 29 печатных трудов.
Имеет 29 печатных трудов.
В 1970 году на базе РКБ одним из первых в стране был создан консультативный кабинет медицинской генетики. Горячим инициатором его создания и становления была заведующая кафедрой нервных болезней БГМИ, профессор Борисова Нинель Андреевна. За короткий промежуток времени, благодаря постоянной и активной поддержке МЗ РБ и администрации больницы были подготовлены квалифицированные кадры медицинских генетиков и организовано оказание медико-генетической помощи населению республики. Параллельно проводились научные исследования сотрудниками кафедры по эпидемиологии и распространенности наследственных заболеваний.
Борисова Н.А. – невролог, доктор медицинских наук, профессор, заслуженный деятель науки БАССР, почетный академик АН РБ, кавалер Ордена Трудового Красного Знамени.
Под ее руководством выполнены и защищены 29 кандидатских диссертаций, является автором более 340 научных трудов, 5 монографий, имеет 14 авторских свидетельств на изобретения. Под ее руководством проведены исследования эпидемиологии, патогенеза, клинических проявлений сирингомиелии, разработаны методы диагностики и лечения, способствующие снижению уровня первичной инвалидности больных.
Под ее руководством проведены исследования эпидемиологии, патогенеза, клинических проявлений сирингомиелии, разработаны методы диагностики и лечения, способствующие снижению уровня первичной инвалидности больных.
Ниннель Андреевна привлекает к себе молодежь, из которой формируются научные и педагогические кадры, руководители отделений, квалифицированные неврологи и генетики, преподаватели кафедры. Первыми преподавателями медицинской генетики студентам БГМИ были БайбазароваФанзияМугалимовна – первая заведующая медико-генетическим кабинетом (1973-1979 гг.), заместитель главного врача РКБ по поликлинике и Флера Габдулхаковна Исхакова–первый педиатр-генетик Республики. Используя свою энергию и целеустремленность, они возглавили работу по своевременному выявлению и лечению врожденных заболеваний обмена веществ у детей, занимались организационной и просветительской работой, читая лекции и проводя занятия с практическими врачами всей республики.
Байбазарова Ф. М. – невролог, кандидат медицинских наук (1985 г.), ассистент кафедры (1979 – 2018 гг.), врач высшей категории, отличник здравоохранения Республики Башкортостан. Трудовую деятельность начала старшим лаборантом кафедры с одновременной учебой в двухгодичной очно-заочно клинической ординатуре. Основные научные исследования посвящены наследственно-дегенеративным заболеваниям нервной системы и перинатальным поражениям нервной системы, детской эпилепсии. Опубликовано 98 научных работ.
М. – невролог, кандидат медицинских наук (1985 г.), ассистент кафедры (1979 – 2018 гг.), врач высшей категории, отличник здравоохранения Республики Башкортостан. Трудовую деятельность начала старшим лаборантом кафедры с одновременной учебой в двухгодичной очно-заочно клинической ординатуре. Основные научные исследования посвящены наследственно-дегенеративным заболеваниям нервной системы и перинатальным поражениям нервной системы, детской эпилепсии. Опубликовано 98 научных работ.
С 1972 года преподавание медицинской генетики для студентов педиатрического и санитарно-гигиенических факультетов проводилось под руководством профессора Мифтаховой А.С., д.м.н. (1973 г.), отличника здравоохранения, главного детского невропатолога МЗ РБ, отличника здравоохранения. Научные исследования Амины Садыковны посвящены изучению вопросов лечения полиомиелита, туберкулезного менингита, детского церебрального паралича. Предложено использование лечебных факторов курорта Красноусольский для комплексного лечения вялых и спастических параличей. Внесла вклад в борьбу с эпидемической вспышкой полиомиелита в республике. Автор 112 научных трудов. Под ее руководством защищены 3 кандидатские диссертации. Награждена медалями.
Внесла вклад в борьбу с эпидемической вспышкой полиомиелита в республике. Автор 112 научных трудов. Под ее руководством защищены 3 кандидатские диссертации. Награждена медалями.
Мифтахова А.С. — невролог, доктор медицинских наук (1973 г.), профессор (1974 г.), отличник здравоохранения г. Уфы. С 1950 г. в БГМИ на кафедре нервных болезней, с 1964 г. — доцент курса физиотерапии, с 1972 г. — заведующая кафедрой нервных болезней педиатрического и санитарно-гигиенического факультетов, с 1982 г. — профессор кафедры неврологии. Одновременно являлась главным физиотерапевтом, главным детским невропатологом МЗ РБ. Научные исследования посвящены изучению вопросов лечения полиомиелита, туберкулезного менингита, детского церебрального паралича.
Развитие и становление медико-генетической службы в РБ проходило под постоянным методическим руководством профессорско-преподавательского состава кафедры неврологии с курсами нейрохирургии и медицинской генетики БГМИ.
С 1989 года кафедру возглавляет доктор медицинских наук, профессор, заслуженный врач Республики Башкортостан и Российской Федерации, кавалер Ордена «За заслуги перед Республикой Башкортостан» Магжанов Рим Валеевич. Одновременно он обеспечивает руководство неврологической и медико-генетической службами республики, будучи главным внештатным неврологом и специалистом по медицинской генетике МЗ РБ (1989 – 2012 гг.).
Магжанов Р.В.- невролог, генетик, доктор медицинских наук (1989 г.), профессор (1991 г.), Заслуженный врач РБ (1997 г.), РФ (2005г.). Докторская диссертация Рима Валеевича посвящена изучению клиники, эпидемиологии, распространенности наследственных нервно-мышечных заболеваний в РБ. Впервые в России им проведено исследование особенностей течения нейро-дегенеративных заболеваний в масштабе крупного региона. Участвовал в выполнении комплексной научной работы по изучению окружающей среды «Здоровье населения Северо-Восточного региона РБ (по программе ЭНТАС АН РБ), которая отмечена Золотым дипломом Международного форума по проблемам науки, техники и образования (М. , 1998 г.). Автор свыше 400 научных работ, в т.ч. 5 монографий, руководства для врачей «Нервные болезни» (М., 2000). Под руководством Р.В. Магжанова защищены 2 докторские и 21 кандидатская диссертации. Член Президиума Правления российского общества неврологов, член Научного совета РАМН и Минздравсоцразвития РФ по неврологии, член Правления РОМГ.
, 1998 г.). Автор свыше 400 научных работ, в т.ч. 5 монографий, руководства для врачей «Нервные болезни» (М., 2000). Под руководством Р.В. Магжанова защищены 2 докторские и 21 кандидатская диссертации. Член Президиума Правления российского общества неврологов, член Научного совета РАМН и Минздравсоцразвития РФ по неврологии, член Правления РОМГ.
Основными направлениями научной работы кафедры неврологии с курсами нейрохирургии и медицинской генетики под руководством Магжанова Р.В. определены: этиология, патогенез, территориально-этническое распространение, клинические проявления и течение наследственных и мультифакторных заболеваний нервной системы. Нейрогенетические исследования вступили с 1993 года в новую фазу своего развития благодаря тесному сотрудничеству с учеными Института биохимии и генетики Уфимского федерального центра РАН под руководством профессора Э.К. Хуснутдиновой. Использование современных молекулярно-генетических технологий распознавания наследственной и наследственно предрасположенной патологии позволило определить спектр и частоту мутаций при наследственной патологии: фенилкетонурия (к. м.н.С.Ш.Мурзабаева), миодистрофия Дюшенна (к.б.н. О.Гринчук), болезнь Вильсона (к.м.н. А.С.Карунас, к.м.н. А.Р.Магжанова), болезнь Шарко-Мари-Тута (к.м.н. Н.Б.Крупина, д.б.н. И.М. Хидиятова), спастические параплегии (к.б.н. Ахметгалеева А.Ф., д.б.н. Хидиятова И.М.), муковисцидоз (д.б.н. Корытина Г.Ф.), болезнь Паркинсона (к.м.н. Байтимиров А.Р., к.б.н. Гилязова И.Р., к.м.н. Ахмадеева Г.И.). Благодаря активному сотрудничеству впервые выявлены особенности клинических и биохимических проявлений, клинико-молекулярно-генетические корреляции при болезнях экспансии тринуклеотидных повторов: миотонической дистрофии (д.м.н. Л.Р.Ахмадеева., д.б.н. Р.И.Фатхлисламова, к.м.н. Е.В.Сайфуллина, к.м.н. В.Ф.Туник, к.м.н. Р.Р.Мухаметова), хорее Гентингтона (к.м.н. И.А.Кутуев), спиноцеребеллярных атаксиях (к.м.н. Э.З.Мингазова). Результаты многолетних совместных научных исследований были обобщены в коллективных монографиях «ДНК-диагностика и профилактика наследственной патологии в Республике Башкортостан» (Уфа, 2005), «Этногеномика народов Центральной Азии: структура генофонда и молекулярно-генетические основы наследственной патологии» под редакцией проф. Э.К.Хуснутдиновой (Уфа,2014).
м.н.С.Ш.Мурзабаева), миодистрофия Дюшенна (к.б.н. О.Гринчук), болезнь Вильсона (к.м.н. А.С.Карунас, к.м.н. А.Р.Магжанова), болезнь Шарко-Мари-Тута (к.м.н. Н.Б.Крупина, д.б.н. И.М. Хидиятова), спастические параплегии (к.б.н. Ахметгалеева А.Ф., д.б.н. Хидиятова И.М.), муковисцидоз (д.б.н. Корытина Г.Ф.), болезнь Паркинсона (к.м.н. Байтимиров А.Р., к.б.н. Гилязова И.Р., к.м.н. Ахмадеева Г.И.). Благодаря активному сотрудничеству впервые выявлены особенности клинических и биохимических проявлений, клинико-молекулярно-генетические корреляции при болезнях экспансии тринуклеотидных повторов: миотонической дистрофии (д.м.н. Л.Р.Ахмадеева., д.б.н. Р.И.Фатхлисламова, к.м.н. Е.В.Сайфуллина, к.м.н. В.Ф.Туник, к.м.н. Р.Р.Мухаметова), хорее Гентингтона (к.м.н. И.А.Кутуев), спиноцеребеллярных атаксиях (к.м.н. Э.З.Мингазова). Результаты многолетних совместных научных исследований были обобщены в коллективных монографиях «ДНК-диагностика и профилактика наследственной патологии в Республике Башкортостан» (Уфа, 2005), «Этногеномика народов Центральной Азии: структура генофонда и молекулярно-генетические основы наследственной патологии» под редакцией проф. Э.К.Хуснутдиновой (Уфа,2014).
По инициативе преподавателей кафедры неврологии с курсами нейрохирургии и медицинской генетики разработаны и внедрены в практику работы МГК компьютерные регистры по моногенным и мультифакторным заболеваниям (хорея Гентингтона, миотоническая дистрофия, фенилкетонурия, болезнь Шарко-Мари-Тута, миастения, болезнь Паркинсона, болезнь Вильсона-Коновалова), которые позволяют упорядочить, систематизировать и анализировать данные по больным, оптимизировать диагностику, диспансерное наблюдение и медико-генетическое консультирование в отягощенных семьях, повысить эффективность профилактических программ в республике. Как признание достижений ученых и медицинских генетиков Башкортостана в г. Уфа в мае 2005 года в соответствии с приказом МЗ РФ проведен V Всероссийский съезд медицинских генетиков с международным участием.
Результаты научных исследований профессорско-преподавательского состава кафедры используются в учебном процессе по курсу Медицинской генетики. К преподаванию медицинской генетики, помимо клиницистов, прошедших подготовку по медицинской генетики, асс., к.м.н., Ф.М. Байбазаровой, доц., к.м.н. У.Б. Давлетшина, доц., д.м.н. С.Ш. Мурзабаевой, доц., к.м.н., А.К. Мардановой, и доц., к.м.н., Е.В. Сайфуллиной привлекались научные сотрудники Института биохимии и генетики УНЦ РАН: член-корр. РАО, профессор Э.К. Хуснутдинова, д.м.н. А.С. Карунас, д.б.н. И.М. Хидиятова.
К преподаванию медицинской генетики, помимо клиницистов, прошедших подготовку по медицинской генетики, асс., к.м.н., Ф.М. Байбазаровой, доц., к.м.н. У.Б. Давлетшина, доц., д.м.н. С.Ш. Мурзабаевой, доц., к.м.н., А.К. Мардановой, и доц., к.м.н., Е.В. Сайфуллиной привлекались научные сотрудники Института биохимии и генетики УНЦ РАН: член-корр. РАО, профессор Э.К. Хуснутдинова, д.м.н. А.С. Карунас, д.б.н. И.М. Хидиятова.
Неоценимый вклад сотрудники кафедры неврологии с курсами нейрохирургии и медицинской генетики внесли в практическое здравоохранение Башкортостана.
Мурзабаева С.Ш. – педиатр, генетик, доктор медицинских наук (2010 г.), Заслуженный врач РФ и РБ, Отличник здравоохранения РФ. Профессор кафедры Салия Шарифьяновна, курирующая в должности заместителя министра здравоохранения РБ службу охраны здоровья матери и ребенка (1998-2007), совмещала управленческую работу с научно-педагогической, внося личный вклад в совершенствование медико-генетической помощи населению республики. В период исполнения полномочий депутата Государственной Думы ФС РФ (2007-2016) продолжала педагогическую деятельность на кафедре.
В период исполнения полномочий депутата Государственной Думы ФС РФ (2007-2016) продолжала педагогическую деятельность на кафедре.
В целях непрерывного профессионального образования врачей республики сотрудниками кафедры регулярно проводятся научно-практические конференции, издаются руководства, методические рекомендации, информационные письма. Кафедра имеет широкие научные связи: международные, с профильными кафедрами ВУЗов РФ, НИИ стран СНГ, с УНЦ РАН. Таким образом, на протяжении 70 лет коллектив кафедры неврологии с курсами нейрохирургии и медицинской генетики проводил плодотворную педагогическую, научно-исследовательскую, лечебно-консультативную и огромную организационно-методическую работу, формируя и возглавляя неврологическую, нейрохирургическую и медико-генетическую службы республики, превратившиеся ныне в крупные научно-практические центры.
В 2018 году по приказу ректора БГМУ, профессора Павлова В.Н., проведена реорганизация кафедры с выделением из ее состава курса медицинской генетики и нейрохирургии. Создана кафедра медицинской генетики и фундаментальной медицины ИДПО БГМУ, которую возглавила д.б.н., профессор, член-корреспондент РАО Эльза Камилевна Хуснутдинова.
Создана кафедра медицинской генетики и фундаментальной медицины ИДПО БГМУ, которую возглавила д.б.н., профессор, член-корреспондент РАО Эльза Камилевна Хуснутдинова.
Хуснутдинова Э.К. – д.б.н., профессор, академик АН РБ, член-корреспондент РАО. Под ее руководством подготовлено 13 докторов, 96 кандидатов наук. Основные направления ее научной деятельности — генетика человека, популяционная генетика, молекулярная генетика. Является Председателем Башкирского отделения РОМГ с 2012 года. Награждена Орденом Дружбы народов Республики Башкортостан (2014 г.), Лауреат Государственной премии РБ в области науки и техники, Заслуженный деятель науки Российской Федерации.
Учебно-педагогический процесс, лечебно-консультативная и организационно-методическая работа сотрудников кафедры ведется на клинических базах: ГБУЗ Республиканский медико-генетический центра (директор, к.б.н., И.Р. Минниахметов), ГБУЗ РБ «Дом ребенка специализированный» города Уфа (гл. врач, Р.Р.Кадикова), Института биохимии и генетики Уфимского федерального исследовательского центра РАН (директор, д.б.н., проф. Э.К. Хуснутдинова).
врач, Р.Р.Кадикова), Института биохимии и генетики Уфимского федерального исследовательского центра РАН (директор, д.б.н., проф. Э.К. Хуснутдинова).
Лекции и практические занятия на кафедре проводятся с использованием современных демонстрационных материалов с помощью мультимедийных презентаций, видеотехники.На кафедре осуществляется преподавание дисциплины «Медицинская генетика» студентам 4 курса лечебного, педиатрического и медико-профилактического факультета. Преподавание осуществляется согласно ГОС. На кафедре преподают 3 профессора, 3 доцента и 3 ассистента. Созданы учебные пособия, электронные и контролирующие программы, видео- и аудиофильмы, подготовлены методические рекомендации для студентов, преподавателей, самостоятельной работы студентов, имеются тестовые задания. Сотрудники кафедры активно принимают участие в работе международных, всероссийских и республиканских конференциях.
На кафедре медицинской генетики и фундаментальной медицины ИДПО проводится обучение по следующим специальностям и дисциплинам:
1. 31.05.01 Лечебное дело – «Медицинская генетика»;
31.05.01 Лечебное дело – «Медицинская генетика»;
2. 31.05.02 Педиатрия – «Медицинская генетика»;
3. 32.05.01 Медико-профилактическое дело – «Медицинская генетика».
Проводится подготовка к лицензированию клинической ординатуры по специальности «Генетика».
Весь учебный процесс на кафедре обеспечен полным набором учебно-методической документации. Учебно-методические материалы (УММ) в размещены на учебном портале БГМУ.
УММ включает:
— методические разработки лекций;
— методические рекомендации для преподавателей;
— методические указания для студентов к практическим занятиям;
— методические указания к самостоятельной аудиторной работе;
— методические указания к самостоятельной внеаудиторной работе;
— комплект тестовых заданий по дисциплине;
— рейтинговая система оценки знаний студентов (критерии оценки знаний и умений студентов по основной дисциплине).
Перечень диссертаций, защищенных профессорско-преподавательским составом кафедры:
1. Хуснутдинова Э.К.: кандидатская диссертация на тему: «Детерминация зачатков разных органов у дрозофилы под действием гомеозисных генов» (1981 г.). Докторская диссертация на тему: «Молекулярно-генетическая характеристика популяций башкир и других народов Волго-Уральского региона» (1997 г.).
Под руководством Хуснутдиновой Э.К. изучена структура генофонда и генетической истории народов Волго-Уральского региона, Средней Азии и Кавказа на основе данных о полиморфизме мт ДНК, У хромосомы, полногеномного анализа 610 000 полиморфных локусов и полногеномного секвенирования. Проведен анализ структурных особенностей генов 15 наследственных и 17 многофакторных заболеваний в различных этнических группах; для ряда популяций Волго-Уральского региона разработаны и успешно применяются алгоритмы ДНК-диагностики наследственных и социально значимых заболеваний. Основные результаты по изучению наследственных заболеваний изложены ею в монографиях “ДНК-диагностика и профилактика наследственной патологии в Республике Башкортостан” и “Этногеномика народов центральной Евразии: структура генофонда и молекулярно-генетические основы наследственной патологии”.
Научные исследования Хуснутдиновой Э.К. поддерживаются международными и российскими фондами: INTAS, Евросоюзом (5 грантов шестой и 7 рамочной программ – GABRIEL, GENOMOS, ADAMS, BCAC, MEDIGENE), Foundation of Alexander von Humboldt, Президиумом РАН (“Фундаментальные науки-медицине” и “Динамика генофондов”), РФФИ, РГНФ, ФЦП «Научные и научно-педагогические кадры инновационной России»; выполняются в сотрудничестве с Центром молекулярной медицины и генетики в Калифорнии (Ирвайн), с Эстонским биоцентром (Тарту), Институтом молекулярной биологии им. М.Планка в Германии (Геттинген), Институтом молекулярной генетики во Франции (Лион) и ERASMUS Университетом в Роттердаме (Нидерланды).
Важнейшие публикации:
— Moffatt M.F., Gut I.G.,…Khusnutdinova E.K., …Demenais F. A large-scale, consortium-based genomewide association study of asthma. NEnglJMed.2010. V. 363(13). P. 1211-21. ИФ – 59,558.
— Pagani L., Lawson D.J., Jagoda E., …, Karunas A., Litvinov S., Khusainova R., Ekomasova N., Akhmetova V., Khidiyatova I., …, Khusnutdinova E.K., Metspalu E., Thomas M.G., Manica A., Nielsen R., Villems R., Willerslev E., Kivisild T., Metspalu M. Genomic analyses inform on migration events during the peopling of Eurasia // Nature. – 2016. – V.538(7624). – P. 238-242. doi: 10.1038/nature19792. ИФ- 38,138.
— Kazantseva A., Gaysina D., Khusnutdinova E. The role of dopamine transporter and dopamine D2 receptor/ankyrin repeat and kinase domain containing 1 gene polymorphisms in personality traits. Prog. NeuropsychopharmacolBiol Psychiatry. 2011. V. 35(4). P.1033-1040. ИФ – 4,025
2. Мурзабаева С.Ш.: кандидатская диссертация на тему: «Клинико-эпидемиологическое и молекулярно-генетическое изучение фенилкетонурии в Республике Башкортостан» (1997 г.). Докторская диссертация на тему: «Оптимизация медико-генетической службы Республики Башкортостан» (2010 г.).
Мурзабаева С.Ш.: кандидатская диссертация на тему: «Клинико-эпидемиологическое и молекулярно-генетическое изучение фенилкетонурии в Республике Башкортостан» (1997 г.). Докторская диссертация на тему: «Оптимизация медико-генетической службы Республики Башкортостан» (2010 г.).
Профессор Мурзабаева С.Ш. занимается вопросами организации медико-генетической помощи населению РФ и РБ, эпидемиологией, клинико-генетическими особенностями распространения наследственных болезней обмена веществ, эффективностью профилактических программ в медицинской генетике.
3. Лукманова Г.И.: кандидатская диссертация на тему: «Иммунотропные свойства бифена и промежуточных продуктов его синтеза» (1996 г.). Докторская диссертация на тему: «Идентификация штаммов Echinococcus granulosus генетические факторы эхинококкоза у детей» (2008 г.).
Профессор Лукманова Г.И принимала участие в исследованиях роли генетических факторов в развитии паразитарных заболеваний человека, в выявлении генотипов возбудителя тяжелого гельминтоза человека – эхинококкоза. С результатами исследований участвует на международных, всероссийских конференциях и форумах. Гульнур Ишмурзовной получено 8 патентов РФ на изобретение.
С результатами исследований участвует на международных, всероссийских конференциях и форумах. Гульнур Ишмурзовной получено 8 патентов РФ на изобретение.
Важнейшие публикации
— СПОСОБ ИДЕНТИФИКАЦИИ ГЕНОТИПА G1 У ИЗОЛЯТОВ ECHINOCOCCUS GRANULOSUS / Буйдаков В.М., Аслаев А.Н., Гумеров А.А., Туйгунов М.М., Лукманова Г.И. / Медицинская паразитология и паразитарные болезни. 2010. № 3. С. 3-5.
— GENETIC DIVERSITY OF ECHINOCOCCUS SPP. IN RUSSIA
Konyaev S.V., Abramov S.A., Krivopalov A.V., Karpenko S.V., Lopatina N.V., Dupal T.A., Yanagida T., Nakao M., Sako Y., Ito A., Ingovatova G.M., Shoykhet Y.N., Bondarev A.Y., Odnokurtsev V.A., Loskutova K.S., Lukmanova G.I., Dokuchaev N.E., Spiridonov S., Alshinecky M.V., Sivkova T.N. et al. / Parasitology. 2013. Т. 140. № 13. С. 1637-1647
— СБОРНИК ЗАДАЧ ПО МЕДИЦИНСКОЙ ГЕНЕТИКЕ И БИОЛОГИИ
учебное пособие для студентов под ред. Викторовой Т.В. / Уфа, 2015.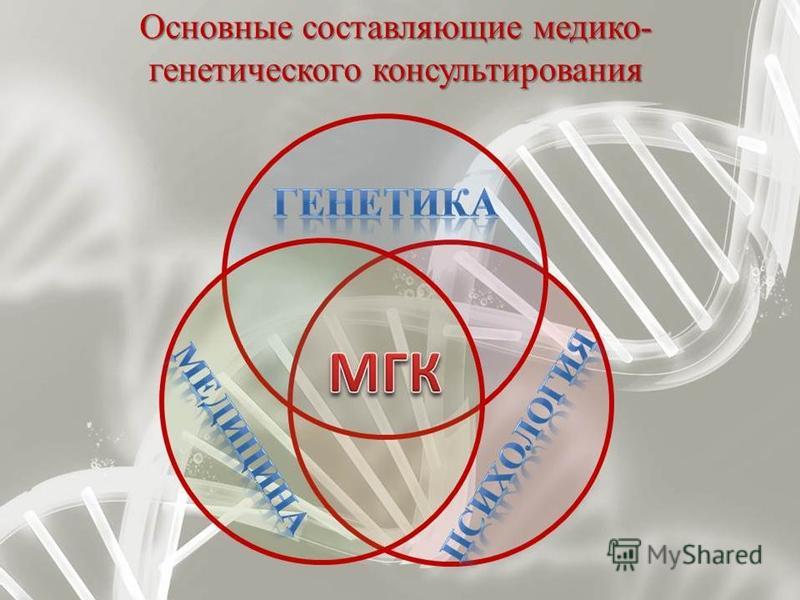 (2-е издание, исправленное и дополненное)
(2-е издание, исправленное и дополненное)
4. Сайфуллина Е.В.: кандидатская диссертация «Структурно-функциональные особенности мембран эритроцитов у больных миотонической дистрофией» (2000 г.).
Доцент Сайфуллина Е.В. изучает территориальные особенности этиологии, клиники наследственных нервно- мышечных заболеваний, спектр мутаций в популяции РБ при данной группе заболеваний.
5. Марданова А.К.: Кандидатская диссертация на тему: «Генетический мониторинг врожденных пороков развития в Республике Башкортостан» (2009г.).
Доцент Марданова А.К. проводит оценку результатов мониторинга ВПР по РБ в сравнении с регистрами субъектов РФ с последующей разработкой профилактических мероприятий в республике.
6. Пушкарева А.Э.: кандидатская диссертация: «Изучение ДНК полиморфизма у молодых больных инфарктом миокарда» (1998г.).
Доцент Пушкарева А.Э. занимается изучением молекулярно-генетических основ различных типов ремоделирования миокарда при сердечной недостаточности, выявлением клинико-генетических моделей прогнозирования исходов при сердечной недостаточности и фибриляции предсердий.
Важнейшие публикации:
— Alfiya E. Pushkareva ,Olga A. Osipova1 , Nina I. Zhernakova2 , Andrey I. Golovin3 , Olga M. Godlevskaya. Pharmacotherapy for the prevention of the progression of chronic heart failure in elderly patients with coronary heart disease. Drug Invention Today. Vol. 10 Special Issue 4.2018.3456-3460
— Alfiya E. Pushkareva ,Olga A. Osipova1 , Nina I. Zhernakova2 Cardiac mechanisms of chronic heart failure development in patients with ischemic heart disease, postman arcticcardiosclerosis of the elderly. Drug Invention Today. Vol.19. Issue 3.2017. 39-43
— Pushkareva A.E., Khusainova R.I., Khusnutdinova E.K., Valiev R.R., Arutunov G.P. Role of angiotensin-converting enzyme polymorphic variants for different types of heart remodeling in patients with chronic heart failure. Journal of Medical Genetics. 2016; 6:11-19.
— Pushkareva A.E., Khusainova R.I., Khusnutdinova E. K., Valiev R.R. The study of expression of growth factor receptor and structure of transforming growth factor gene in heart failure. International Research Journal.2016; 9 (51) P.3: 69-77.
K., Valiev R.R. The study of expression of growth factor receptor and structure of transforming growth factor gene in heart failure. International Research Journal.2016; 9 (51) P.3: 69-77.
— Pushkareva A.E., Khusainova R.I., Khusnutdinova E.K. Polymorphic variants of angiotensin-converting enzyme for different types of heart remodeling. Materials of 4th International Forum «Russian Days of Heart» 21-23 April 2016, St-Petersburg: 71-72.
— Khusainova R.I., Pushkareva A.E., Valiev R.R., Khusnutdinova E.K. Search of structural changes for Transforming Growth Factor Beta1 Receptor Gene (TGFβR1) in patients with chronic heart failure. Genetics. 2014; V.50, (5): 1-8.
— Pushkareva A.E., Khusainova R.I., Arutunov G.P., Khusnutdinova E.K. Study of polymorphic variants of cytokines genes (TNFA, TNFB) and gene methylenetetrafolatereductase (MTHFR) at different types of heart remodeling. Medical Мessenger of Bashkortostan.2012. 4: 81-84.
— Pushkareva A.E., Khusainova R.I., Khusnutdinova N.N., Khusnutdinova E.K. Genotypes HFE in patients with heart failure: clinic-genetics aspects and prognosis. Journal of Heart Failure. 2011. V.(12).6:350-356
— Khusainova R.I., Lebedeva E.R., Pushkareva A.E., Khusnutdinova E.K. The analysis of polymorphic variants of collagen type III gene (COL3A1) in patients with arterial aneurysms of cerebral vessels. Kursk Scientific and Practical Messenger «Human and his Health». 2011.2:45-51.
— Pushkareva A.E., Enikeeva G.R., Kuchina T.A., Khusainova R.I. The study of expression of growth receptors and troponin C for heart failure. Journal of Heart Failure. 2010; №2:243-248.
7. Тимашева Я.Р.: «Роль полиморфизма и уровня экспрессии генов цитокиновой сети в формировании молекулярно-генетических основ предрасположенности к эссенциальной гипертензии»(2008г.).
Доцент Тимашева Я.Р. занимается изучением молекулярно-генетических основ возникновения сердечно-сосудистых заболеваний, вопросов старения и долголетия.
Важнейшие публикации:
— Y Timasheva, T Nasibullin, V Erdman, I Tuktarova, O Mustafina (2018) CXCL13 polymorphism is associated with essential hypertension in Tatars from Russia. Molecular Biology Reports. https://doi.org/10.1007/s11033-018-4257-x
— Y. Timasheva, T. Nasibullin, E. Imaeva, V. Erdman, I. Tuktarova, O. Mustafina (2017) Genetic determinants of essential hypertension in the population of Tatars from Russia. J Hypertens. 2017. 35:S16-S23. https://doi.org/10.1097/HJH.0000000000001332
— T.R. Nasibullin, *Y.R. Timasheva, R.I. Sadikova, I.A. Tuktarova, V.V. Erdman, I.E. Nikolaeva, O.E. Mustafina (2016) Genotype/allelic combinations as potential predictors of myocardial infarction. Molecular Biology Reports. 43(1): 11-16. https://doi.org/10.1007/s11033-015-3933-3
— Y.R.Timasheva, T.R.Nasibullin, O.E. Mustafina (2015) The CXCR2 gene polymorphism is associated with stroke in patients with essential hypertension. Cerebrovascular Diseases Extra. 5(3): 124-131. https://doi.org/10.1159/000441529
Cerebrovascular Diseases Extra. 5(3): 124-131. https://doi.org/10.1159/000441529
— Y.R. Timasheva, T.R. Nasibullin, E.B. Imaeva, V.V. Erdman, I.A. Tuktarova, I.E. Nikolaeva, O.E. Mustafina (2015) Polymorphisms of inflammatory markers and risk of essential hypertension in Tatars from Russia. Clinical Experimental Hypertension. 37(5): 398-403. https://doi.org/10.3109/10641963.2014.987394
— Y. Timasheva, M. Putku, R. Kivi, V. Kozich, J. Mannik, M. Laan (2013) Developmental programming of growth: Genetic variant in Gh3 gene encoding placental growth hormone contributes to adult height determination. Placenta. 34(11): 995-1001. https://doi.org/10.1016/j.placenta.2013.08.012
— Y. R. Timasheva, T. R. Nasibullin, A. N. Zakirova, O. E. Mustafina (2008) Association of interleukin-6, interleukin-12 and interleukin-10 genes polymorphisms with essential hypertension in Tatars from Russia. Biochemical Genetics. 46 (1-2): 64-74. https://doi. org/10.1007/s10528-007-9130-x
org/10.1007/s10528-007-9130-x
— О.В. Заплахова, Я.Р. Тимашева, К.З. Бахтиярова, И.А. Туктарова, О.Е. Мустафина (2017) Клинический и молекулярно-генетический анализ случая семейного рассеянного склероза в Республике Башкортостан». Журнал неврологии и психиатрии им. С.С. Корсакова. 117(2):31.
— Каримов Д.Д., Эрдман В.В., Насибуллин Т.Р., Туктарова И.А., Сомова Р.Ш., Тимашева Я.Р., Мустафина О.Е. (2016) Alu-инсерционно-делеционный полиморфизм генов COL13A1 и LAMA2: анализ ассоциаций с долгожительством. Генетика. 52(10):1185-1193.
— Насибуллин Т.Р., Ягафарова Л.Ф., Ягафаров И.Р., Тимашева Я.Р., Эрдман В.В., Туктарова И.А., Мустафина О.Е. (2016) Сочетания полиморфных маркеров генов белков острой фазы воспаления, хемокинов и их рецепторов как потенциальные предикторы ишемической болезни сердца Acta Naturae. 1 (28):121-126.
— Я. Р. Тимашева, Т. Р. Насибуллин, И. А. Туктарова, В. В. Эрдман, И. Е. Николаева, О. Е. Мустафина (2015) Анализ ассоциаций полиморфных маркеров хемокинов и хемокиновых рецепторов с эссенциальной гипертензией. Молекулярная медицина. 3: 62-64.
Е. Мустафина (2015) Анализ ассоциаций полиморфных маркеров хемокинов и хемокиновых рецепторов с эссенциальной гипертензией. Молекулярная медицина. 3: 62-64.
— Т. Р. Насибуллин, Я.Р. Тимашева, И.А. Туктарова, В.В. Эрдман, И.Е. Николаева, О.Е. Мустафина (2014) Сочетания полиморфных маркеров генов цитокиновой сети как потенциальные предикторы инфаркта миокарда. Генетика, 50(9), 1116-1123.
— Т.Р. Насибуллин, Р.И. Садикова, Я.Р.Тимашева, И.А. Туктарова, В.В. Эрдман, Л.Н. Хусаинова, И.Е. Николаева, О.Е. Мустафина (2014) Ассоциации полиморфных маркеров генов воспаления с инфарктом миокарда. Генетика, 50(1), 236-242.
— Я. Р. Тимашева, Т. Р. Насибуллин, И.А. Туктарова, И.М. Карамова, А.Н. Петрин, О. Е. Мустафина (2012) Анализ межгенных взаимодействий полиморфных локусов генов цитокинов у больных эссенциальной гипертензией. Артериальная гипертензия. 5 (18): 443-448.
— О. Е. Мустафина, Я. Р. Тимашева, Г. Х. Тулякова, Т. Р. Насибуллин, И. А. Туктарова (2010) Полиморфизм -627С>A гена интерлейкина 10: анализ ассоциаций с сердечно-сосудистыми заболеваниями. Медицинская генетика. 9 (5): 12-17.
Р. Насибуллин, И. А. Туктарова (2010) Полиморфизм -627С>A гена интерлейкина 10: анализ ассоциаций с сердечно-сосудистыми заболеваниями. Медицинская генетика. 9 (5): 12-17.
— О. Е. Мустафина, Я. Р. Тимашева (2008). Цитокины и атеросклероз: молекулярные механизмы патогенеза. Молекулярная медицина, № 1, С. 56-68.
— Я. Р. Тимашева, Т. Р. Насибуллин, А. Н. Закирова, О. Е. Мустафина (2007). Полиморфизм гена фактора некроза опухолей альфа у больных эссенциальной гипертензией. Кардиоваскулярная терапия и профилактика, № 6, С. 5-9.
— Я. Р. Тимашева, Г. Х. Тулякова, Т. Р. Насибуллин, А. Н. Закирова, И. М. Карамова, О. Е. Мустафина (2008) Анализ полиморфного варианта -627С/А гена интерлейкина-10 у больных сердечно-сосудистыми заболеваниями Республики Башкортостан. Уральский медицинский журнал. № 7(35), С. 68-71.
— Я.Р. Тимашева, Д.Х. Валиахметова (2001) Полиморфизм генов цитокинов и их рецепторов в популяциях Волго-Уральского региона. Здравоохранение Башкортостана, № 8, С. 55-57.
Здравоохранение Башкортостана, № 8, С. 55-57.
8. Мустафин Р.Н.: «Эпидемиологическое и молекулярно-генетическое исследование нейрофиброматоза I типа в Республике Башкортостан»(2012г.).
Доцент Мустафин Р.Н. проводит поиск мутаций в гене нейрофиброматоза 1-го типа у больных из Республики Башкортостан, занимается вопросами онкогенетики, психогенетики, эпигенетики и эволюционной генетики.
Важнейшие публикации
— R.N. Mustafin and E.K. Khusnutdinova. The Role of Transposable Elements in Emergence of Metazoa // Biochemistry (Moscow). 2018. V. 83. № 3. P. 185-199. https://www.ncbi.nlm.nih.gov/pubmed/29625540
— R.N. Mustafin and E.K. Khusnutdinova. The Role of Transposons in Epigenetic Regulation of Ontogenesis // Russian Journal of Developmental Biology. 2018. V. 49. №2. P. 61-78. https://elibrary.ru/show_item_pages.asp?fname=elibrary_32675476_74810769
— Mustafin R. N., E.K. Khusnutdinova. Epigenetic Hypothesis of the Role of Peptides in Aging // Advances in Gerontology. 2018. V. 8. No. 3. pp. 200-209. https://www.ncbi.nlm.nih.gov/pubmed/29860723
N., E.K. Khusnutdinova. Epigenetic Hypothesis of the Role of Peptides in Aging // Advances in Gerontology. 2018. V. 8. No. 3. pp. 200-209. https://www.ncbi.nlm.nih.gov/pubmed/29860723
— Mustafin R.N., Khusnutdinova E.K. The role of interactions of transposons with epigenetic factors in the aging process // Adv. Gerontol. 2017. V. 30. N 4. P. 516-528. https://www.ncbi.nlm.nih.gov/pubmed/28968025
— Mustafin R.N., Khusnutdinova E.K. The Role of Transposable Elements in Ontogenesis // Usp. Fiziol. Nauk. 2016. V. 47. N 3. P. 70-96. https://www.ncbi.nlm.nih.gov/pubmed/29283231
— Mustafin R.N. The Role of Viruses in the Genome Evolution // Usp. Fiziol. Nauk. 2016. V. 47. N 2. P. 77-97. https://www.ncbi.nlm.nih.gov/pubmed/27530045
— Mustafin R.N., Khusnutdinova E.K. The role of introns in evolution // Usp. Fiziol. Nauk. 2015. V. 46. N 1. P. 93-104. https://www.ncbi.nlm.nih.gov/pubmed/25854116
— Мустафин Р. Н., Хуснутдинова Э.К. Общие универсальные генетические закономерности, лежащие в основе возникновения и эволюции многоклеточности // Успехи физиологических наук. 2018. Т. 49. № 3. С. 52-81. https://elibrary.ru/projects/pdf_viewer/pdf_viewer.asp?fileid=35324266
Н., Хуснутдинова Э.К. Общие универсальные генетические закономерности, лежащие в основе возникновения и эволюции многоклеточности // Успехи физиологических наук. 2018. Т. 49. № 3. С. 52-81. https://elibrary.ru/projects/pdf_viewer/pdf_viewer.asp?fileid=35324266
— Мустафин Р.Н. Особенности эпигенетической регуляции онтогенеза растений // Успехи современной биологии. 2018. № 3. С. 227-242. https://elibrary.ru/projects/pdf_viewer/pdf_viewer.asp?fileid=35324452
— Мустафин Р.Н., Хуснутдинова Э.К. Взаимосвязь прионов с некодирующими РНК // Вавиловский журнал генетики и селекции. 2018. Т. 22. № 4. С. 856-865. http://www.bionet.nsc.ru/vogis/download/06_Mustafin.pdf
— Мустафин Р.Н., Хуснутдинова Э.К. Роль транспозонов в возникновении многоклеточных животных // Биохимия. 2018. T 83. Вып. 3. С. 291-308.
— Мустафин Р.Н., Хуснутдинова Э.К. Роль транспозонов в эпигенетической регуляции онтогенеза // Онтогенез. 2018. № 2. С. 61-78.
№ 2. С. 61-78.
— Мустафин Р.Н., Хуснутдинова Э.К. Эпигенетическая гипотеза пептидной регуляции старения // Успехи геронтологии. 2018. Т. 31. № 1. С. 10-20.
— Мустафин Р.Н., Хуснутдинова Э.К. Некодирующие части генома как основа эпигенетической наследственности // Вавиловский журнал генетики и селекции. 2017. Т. 21. № 6. С. 742-749. http://www.bionet.nsc.ru/vogis/download/22_Mustafin.pdf
— Мустафин Р.Н., Хуснутдинова Э.К. Роль эпигенетических факторов в патогенезе нейрофиброматоза 1-го типа // Успехи молекулярной онкологии. 2017. № 3. C. 15-27. http://umo.abvpress.ru/jour/issue/archive
— Мустафин Р.Н., Хуснутдинова Э.К. Взаимосвязь эпигенетических факторов в механизмах старения и малигнизации // Успехи физиологических наук. 2017. № 3. С. 92-120. http://i.uran.ru/webcab/journals/journal/uspehi-fiziologicheskih-nauk/uspehi-fiziologicheskih-nauk-2017-t48-n-2
— Мустафин Р.Н. Асептический некроз головки бедренной кости // Лечебное дело. 2015. № 4. С. 7-20. https://elibrary.ru/item.asp?id=25627442
2015. № 4. С. 7-20. https://elibrary.ru/item.asp?id=25627442
— Мустафин Р.Н., Хуснутдинова Э.К. Молекулярные механизмы развития остеоартроза // Лечебное дело. 2015. № 3. С. 86-93. https://elibrary.ru/item.asp?id=25518823
— Мустафин Р.Н., Бермишева М.А., Хуснутдинова Э.К. Особенности нейрофиброматоза 1-го типа в республике Башкортостан // Медицинская генетика. 2015. Т. 14. № 6 (156). С. 29-35. https://elibrary.ru/item.asp?id=24400931
9. Гилязова И.Р.: «Молекулярно-генетическое изучение болезни Паркинсона в Башкортостане» (2004г.).
Доцент Гилязова И.Р. проводит изучение генетических и эпигенетических основ онкоурологических заболеваний.
— Кутлыева Л.Р., Гилязова И.Р., Хусаинова Р.И., Хуснутдинова Э.К. Роль модификации гистонов и метилирования ДНК в развитии почечно-клеточных карцином// Экологическая генетика. — 2012. — Т. 10. — № 3.- С. 59-76.
— Л.Р. Кутлыева, И. Р. Гилязова, Р.И. Хусаинова, А.А. Загидуллин, А.А. Халлиуллин, Е.А. Климентова, А.М. Шафигина, В.Н. Павлов, Э.К. Хуснутдинова Поиск изменений нуклеотидной последовательности в гене фон Хиппеля-Линдау и анализ аллельных делеций в генах-супрессорах опухолевого роста у больных светлоклеточным раком почки из Республики Башкортостан // Медицинская генетика. – 2012. – №12. – С. 27-35.
Р. Гилязова, Р.И. Хусаинова, А.А. Загидуллин, А.А. Халлиуллин, Е.А. Климентова, А.М. Шафигина, В.Н. Павлов, Э.К. Хуснутдинова Поиск изменений нуклеотидной последовательности в гене фон Хиппеля-Линдау и анализ аллельных делеций в генах-супрессорах опухолевого роста у больных светлоклеточным раком почки из Республики Башкортостан // Медицинская генетика. – 2012. – №12. – С. 27-35.
— Ахмадишина Л.З., Гилязова И.Р., Кутлыева Л.Р., Корытина Г.Ф., Кочетова О.В., Урманцев М.Ф., Измайлова С.М., Измайлов А.А., Кунсбаева Г.Б., Загидуллин А.А., Халиуллин А.А., Павлов В.Н., Викторова Т.В., Хуснутдинова Э.К. Ассоциация полиморфных вариантов генов репарации ДНК XRCC1, XPD с развитием рака мочевого пузыря и рака почки// Генетика. — 2014. — Т. 50.- № 4. — С. 481-490.
— Гилязова И.Р., Кунсбаева Г.Б., Янкина М.А., Мустафин А.Т., Сафиуллин Р.И., Павлов В.Н., Хуснутдинова Э.К. Герминальные мутации в генах системы репарации ДНК у пациентов с раком предстательной железы// Медицинская генетика. — 2016. — Т. 15. — № 7 (169). — С. 21-23.
— 2016. — Т. 15. — № 7 (169). — С. 21-23.
— Кунсбаева Г.Б., Гилязова И.Р., Янкина М.А., Климентова Е.А., Павлов В.Н., Хуснутдинова Э.К. Поиск мутаций в гене синдрома Блума (BLM) у пациентов с раком предстательной железы// Медицинская генетика. 2016. Т. 15. № 5 (167). С. 7-9.
— Гилязова И.Р. Янкина М.А., Кунсбаева Г.Б., Климентова Е.А., Измайлов А.А., Павлов В.Н., Хуснутдинова Э.К. Характеристика мутаций в опухолях предстательной железы на основании данных секвенирования экзома // Генетика. – 2016. – Т. 52. – №. 9. С.1097-1103.
— Климентова Е.А., Гилязова И.Р., Кунсбаева Г.Б., Измайлов А.А., Султанов И.М., Павлов В.Н., Хуснутдинова Э.К. Ассоциация полиморфного варианта сайта связывания микроРНК rs10491534 гена TSC1 с тяжестью течения рака почки // Медицинская генетика. – 2016. – T.15. – №4. – С.50-52.
— Климентова Е.А., Гилязова И.Р., Измайлов А.А., Султанов И.М., Павлов В.Н., Хуснутдинова Э.К. Идентификация мутаций в гене ремоделирования хроматина PBRM1 у пациентов со светлоклеточным раком почки// Вавиловский журнал генетики и селекции. – 2018. – №22(7) – С.873-877.
– 2018. – №22(7) – С.873-877.
— Бермишева М.А., Тахирова З.Р., Гилязова И.Р., Хуснутдинова Э.К. Ассоциация полиморфных вариантов генов биогенеза микроРНК с развитием рака молочной железы// Генетика. – 2018. – Т. 54. – № 5. – С. 563-572.
— Mavaddat N, Michailidou K, Dennis J,….Bermisheva M.A., …. Gilyazova I.R., ….. Khusnutdinova E.K Polygenic Risk Scores for Prediction of Breast Cancer and Breast Cancer Subtypes // Am J Hum Genet. – 2019 – 104(1) – P.21-34.
На кафедре медицинской генетики и фундаментальной медицины ИДПО БГМУ организован студенческий научный кружок (https://vk.com/geneticabashgmu), ответственный по кафедре – к.б.н., доцент Мустафин Р.Н. В течение года проведено 8 заседаний, с активным участием 12-15 студентов. Темы заседаний кружка выбраны с учетом актуальности проблемы и интереса студенческой молодежи. Презентации студентов – участников кружка были посвящены вопросам диагностики хромосомных заболеваний, моногенных наследственных болезней обмена веществ, применения методов медицинской статистики в клинической генетике, профилактике наследственных болезней, обзору актуальных методов лечения орфанных заболеваний. Проведено совместное заседание СНК кафедры Госпитальной Терапии 2 (зав.каф.: Бакиров Булат Ахатович), посвященное генетическим основа остеоартроза и асептического некроза головки бедренной кости.
Проведено совместное заседание СНК кафедры Госпитальной Терапии 2 (зав.каф.: Бакиров Булат Ахатович), посвященное генетическим основа остеоартроза и асептического некроза головки бедренной кости.
В текущем учебном году запланировано проведение заседаний с участием преподавателей кафедры с учетом направлений их научной деятельности: скринирующие программы в РБ, эффективность, перспективы; энзимопатии, патогенетическая терапия при метаболических нарушениях.
ОТЧЕТ ПО НИР
Краевая медико — генетическая консультация
Воронин Сергей Владимирович
Заведующий отделением КМГК, кандидат медицинских наук,главный специалист МЗ РФ по медицинской генетике в Дальневосточном Федеральном округе, врач высшей категории.
Информационные статьи:
Краевая Медико Генетическая консультация (КМГК), оказывающая помощь пациентам и супружеским парам, работает в Приморском крае с 1976 года, в том числе, как подразделение ККЦСВМП с 1996 года.
Мы являемся современной, высокотехнологической службой, способной оказать помощь Вам или членам Вашей семьи по широкому кругу вопросов и проблем связанным с наследственной патологией, врожденными пороками развития, мультифакториальными болезнями, планированием «оптимального» ребенка, бесплодием, невынашиванием беременности и др.
- Отделение медико-генетического консультирования
- Кабинет ультразвуковой диагностики плода
- Отделение пренатальной диагностики
- Лаборатория гормональных исследований
- Лаборатория цитогенетики
- Лаборатории селективного скрининга наследственных заболеваний
- Лаборатории массового скрининга наследственных заболеваний
- Лаборатория молекулярно-генетических исследований
В этих подразделениях на современном оборудовании работают сертифицированные специалисты, прошедшие обучение в ведущих клиниках и научно-исследовательских институтах России.
Мы рады оказать Вам следующие виды медицинской помощи:
Отделение медико генетического консультирования (время работы с 8.00 до 20.00):
- Проведут планирование “оптимального” ребенка
- Составят и выполнят план обследования беременной женщины на наследственную патологию у плода
- Окажут помощь при решении вопросов по вынашиванию беременности или прерыванию ее по медицинским показателям
- Окажут квалифицированную помощь при различных психологических нарушениях
- Составят родословную карту семьи
- Уточнят природу заболевания (наследственное, ненаследственное, мультифакториальное)
- Сделают фенотипическое описание
- Определят риск развития заболеваний (рак, гемофилия, миопатии, дальтонизм и др.)
- Дадут прогноз здоровья потомства
- Проконсультируют по вопросам установления отцовства
- Если Вам необходимо, возможно прочтение лекций по вопросам медицинской генетики в Вашем учреждении
В этом отделении для Вас работают:
Воронин Сергей Владимирович — заведующий КМГК, кандидат медицинских наук, главный специалист МЗ РФ по медицинской генетике в Дальневосточном Федеральном округе, врач высшей категории.
Воронина Валентина Георгиевна — к.б.н., зав. консультативным отделением с пренатальной диагностикой КМГК, врач высшей категории.
Пронина Елена Николаевна — врач-генетик, врач второй категории.
Милкина Екатерина Юрьевна — врач-генетик, врач второй категории.
Сергеева Наталья Юрьевна — врач-генетик, врач второй категории.
Михайлик Лариса Ивановна — врач-генетик.
Решетова Ольга Александровна – медицинский психолог, специалист первой категории.
Кабинет ультразвуковой диагностики плода:
Только у нас в Приморском крае проводится УЗИ плода 2-го уровня, все остальные УЗИ плода, даже проведенные в учреждении, имеющем аппаратуру и специалистов экспертного класса, все равно являются УЗИ плода 1-го уровня.
В случае выявления патологии у плода (или подозрения на нее) на 1-м уровне для подтверждения или снятия заключения Вам будет необходимо проведение УЗИ плода в КМГК.
В этом отделении для Вас работают:
- Капацинская Оксана Владимировна — врач ультразвуковой диагностики, врач акушер-гинеколог по пренатальной диагностике, к.м.н., врач высшей категории
- Горлов Роман Александрович – врач ультразвуковой диагностики, акушер-гинеколог
- Дударь Оксана Анатольевна — врач ультразвуковой диагностики, врач акушер-гинеколог по пренатальной диагностике, к.м.н., врач высшей категории
- Макарчук Светлана Геннадьевна– врач ультразвуковой диагностики, акушер-гинеколог, врач высшей категории, врач высшей категории
- Василенко Елена Михайловна – врач ультразвуковой диагностики, акушер-гинеколог, врач высшей категории
- Полинтова Ольга Николаевна – врач ультразвуковой диагностики, акушер-гинеколог, врач высшей категории
Отделение инвазивной пренатальной диагностики:
- Выполнят генетическое исследование плода с целью предупреждения рождения ребенка с хромосомной патологией (болезнь Дауна, Шерешевского-Тернера и др.
 )
) - Установят пол плода с 9 недели беременности
- Определят группу крови и резус крови плода с 21 недели беременности
- Определят наличие инфекции в крови плода и околоплодных водах
- Проведут коррекцию количества околоплодных вод (при маловодии и многоводии).
- Проведут внутриутробно операции по коррекции некоторых врожденных пороков развития плода
- Проведут забор плодного материала для проведения молекулярных методов диагностики для определения наследственного заболевания у плода
- В том случае, если Вы не уверены, кто является отцом плода, мы можем помочь установить Вам это, начиная с 9-ой недели беременности.
 )
)- Капацинская Оксана Владимировна — врач ультразвуковой диагностики, врач акушер-гинеколог по пренатальной диагностике, к.м.н., врач высшей категории
- Горлов Роман Александрович – врач ультразвуковой диагностики, акушер-гинеколог
- Дударь Оксана Анатольевна — врач ультразвуковой диагностики, врач акушер-гинеколог по пренатальной диагностике, к. м.н., врач высшей категории
- Макарчук Светлана Геннадьевна– врач ультразвуковой диагностики, акушер-гинеколог, врач высшей категории, врач высшей категории
- Василенко Елена Михайловна – врач ультразвуковой диагностики, акушер-гинеколог, врач высшей категории
- Полинтова Ольга Николаевна – врач ультразвуковой диагностики, акушер-гинеколог, врач высшей категории
 м.н., врач высшей категории
м.н., врач высшей категории
Лаборатория гормональных исследований
(время работы 8.00- 16.00, время забора крови 8.00-19.00 рабочие дни кааб. 809, суббота 9.00-14.00, первый этаж, приемное отделение).
На современном высокоточном оборудовании флюорометрическом анализаторе «Autodelfia» и биохимическом анализаторе автоматического типа «AIO-360» с использованием только импортных фирменных реагентов Вам:
- Определят во время беременности содержание в крови гормонов РАРР-А, АФП, ХГЧ, свободного эстриола которые являются маркерами хромосомной патологии (болезнь Дауна и др. ) и врожденных пороков развития плода. С компьютерном расчетом степени риска патологии в программе
- Определят полный гормональный статус пациентов при бесплодии, невынашивании беременности, эндокринных и других заболеваниях.
 ) и врожденных пороков развития плода. С компьютерном расчетом степени риска патологии в программе
) и врожденных пороков развития плода. С компьютерном расчетом степени риска патологии в программеВ этом отделении для Вас работает:
- Бруенок Елена Валерьевна — врач-лаборант-генетик высшей категории.
- Чаусова Полина Александровна – врач лабораторный генетик.
(время работы с 8.00 до16.00)
- Сделают исследование кариотипа (определят хромосомный набор) при множественных врожденных пороках развития, умственной отсталости, нарушениях строения наружных или внутренних половых органов.
- Если у Вас есть бесплодие или нарушение менструального цикла неясной этиологии, невынашивание беременности, рождении ребенка с врожденными пороками развития или умственной отсталостью – Вас необходимо провести исследование кариотипа на носительство сбалансированной перестройки хромосом.
- Если у Вас произошло самопроизвольное прерывание беременности (замершая беременность) Вам необходимо исследовать хромосомный набор замершего плода. Считается, что до 50-60% таких проблем до 12 недель беременности , вызваны нарушением хромосомного набора плода. Если причина самопроизвольного прерывания была действительно в этом – Вам необходимо проконсультироваться у врача-генетика, так как у Вас есть повышенный риск повторения этой проблемы.
- Если есть подозрение об изменении числа половых хромосом (например при синдроме Шерешевского-Тернера или Клайнфельтера) мы можем провести исследование полового хроматина (экспресс диагностика пола).
В этом отделении для Вас работают:
- Шапкина Наталья Николаева- врач лабораторный генетик высшей категории
- Михайлик Лариса Ивановна – врач лабораторный генетик
- Тюрина Ольга Владимировна — биолог
- Михайлова Ирина Евгеньевна — биолог
(время работы с 8. 00 до 16.00)
00 до 16.00)
- Сделают пилокарпииновый и рентгенпленочный тест при подозрении на муковисцидоз (нарушения со стороны пищеварительного тракта, длительные, частые, трудно подающиеся лечения проостудные заболевания и др.)
- Выполнят анализ на альфа-антиприпсин (для исключения ферментативной недостаточности при хронических заболеваниях легких)
- Проведут тонкослойную хроматографию и другие исследования при наследственных болезнях обмена веществ
- Сделают реакции ЦТАБ и ЦПХ при подозрении на дисплазии соединительной ткани (вывихи суставов, сколиоз, переломы, пролапс митрального клапана и др.)
- Реакцию Бенедикта при подозрении на лактазную недостаточность (непереносимость молока)
В этом отделении для Вас работают:
- Чаусова Полина Александровна – врач лабораторный генетик
- Пожарская Ирина Николаевна – врач-лабораторный генетик
- Егорова Светлана Владимировна – врач лабораторный генетик, первая категория
(время работы с 8. 00 до 16.00)
00 до 16.00)
- Бесплатно проведут обследование ребенка на фенилкетонурию, врожденный гипотериоз, муковисцидоз, адреногенитальный синдром, галактоземию (часто встречающиеся наследственные заболевания, которые в отсутствии лечения приводят к умственной отсталости, поражениям внутренних органов вплоть до гибели ребенка)
- Определят гетерозиготное носительство гена фенилкетонурии (его распространенность в Приморье — примерно каждый сороковой житель)
В этом отделении для Вас работает:
-
Бруенок Елена Валерьевна — врач-лаборант генетик 1 категории
-
Чаусова Полина Александровна – врач лабораторный генетик
FISH-диагностика – метод позволяющий в узнать была ли причиной ВПР плода или замершей беременности – хромосомная патология у плода
Лаборатория молекулярно-генетических видов исследования:
- Проводится диагностика моногенных болезней (муковисцидоз, болезнь Вильсона-Коновалова и др. )
- Изучение полиморфизмов в генах предрасположенности к сложнонаследуемым заболеваниям (невынашивание беременности, тромбозы, инфаркты, инсульты, сахарный диабет, рак молочной железы и др.).
- Фармакогенетика – подбор лекарственных препаратов (антикоагулянты) с учетом особенностями генетического паспорта, что позволяет снизить риск осложнений от лечения.
 )
)
В этом отделении для Вас работает:
- Воронин Сергей Владимирович — заведующий КМГК, кандидат медицинских наук, врач лабораторный генетик
- Егорова Светлана Владимировна – врач лабораторный генетик, первая категория
- Чаусова Полина Александровна – врач лабораторный генетик
- Тюрина Ольга Владимировна — биолог
Российский стартап в области исследований ДНК Genotek привлёк $4 млн от «Яндекса» и других инвесторов Статьи редакции
{«id»:190625,»url»:»https:\/\/vc.ru\/services\/190625-rossiyskiy-startap-v-oblasti-issledovaniy-dnk-genotek-privlek-4-mln-ot-yandeksa-i-drugih-investorov»,»title»:»\u0420\u043e\u0441\u0441\u0438\u0439\u0441\u043a\u0438\u0439 \u0441\u0442\u0430\u0440\u0442\u0430\u043f \u0432 \u043e\u0431\u043b\u0430\u0441\u0442\u0438 \u0438\u0441\u0441\u043b\u0435\u0434\u043e\u0432\u0430\u043d\u0438\u0439 \u0414\u041d\u041a Genotek \u043f\u0440\u0438\u0432\u043b\u0451\u043a $4\u00a0\u043c\u043b\u043d \u043e\u0442 \u00ab\u042f\u043d\u0434\u0435\u043a\u0441\u0430\u00bb \u0438 \u0434\u0440\u0443\u0433\u0438\u0445 \u0438\u043d\u0432\u0435\u0441\u0442\u043e\u0440\u043e\u0432″,»services»:{«facebook»:{«url»:»https:\/\/www. facebook.com\/sharer\/sharer.php?u=https:\/\/vc.ru\/services\/190625-rossiyskiy-startap-v-oblasti-issledovaniy-dnk-genotek-privlek-4-mln-ot-yandeksa-i-drugih-investorov»,»short_name»:»FB»,»title»:»Facebook»,»width»:600,»height»:450},»vkontakte»:{«url»:»https:\/\/vk.com\/share.php?url=https:\/\/vc.ru\/services\/190625-rossiyskiy-startap-v-oblasti-issledovaniy-dnk-genotek-privlek-4-mln-ot-yandeksa-i-drugih-investorov&title=\u0420\u043e\u0441\u0441\u0438\u0439\u0441\u043a\u0438\u0439 \u0441\u0442\u0430\u0440\u0442\u0430\u043f \u0432 \u043e\u0431\u043b\u0430\u0441\u0442\u0438 \u0438\u0441\u0441\u043b\u0435\u0434\u043e\u0432\u0430\u043d\u0438\u0439 \u0414\u041d\u041a Genotek \u043f\u0440\u0438\u0432\u043b\u0451\u043a $4\u00a0\u043c\u043b\u043d \u043e\u0442 \u00ab\u042f\u043d\u0434\u0435\u043a\u0441\u0430\u00bb \u0438 \u0434\u0440\u0443\u0433\u0438\u0445 \u0438\u043d\u0432\u0435\u0441\u0442\u043e\u0440\u043e\u0432″,»short_name»:»VK»,»title»:»\u0412\u041a\u043e\u043d\u0442\u0430\u043a\u0442\u0435″,»width»:600,»height»:450},»twitter»:{«url»:»https:\/\/twitter.
facebook.com\/sharer\/sharer.php?u=https:\/\/vc.ru\/services\/190625-rossiyskiy-startap-v-oblasti-issledovaniy-dnk-genotek-privlek-4-mln-ot-yandeksa-i-drugih-investorov»,»short_name»:»FB»,»title»:»Facebook»,»width»:600,»height»:450},»vkontakte»:{«url»:»https:\/\/vk.com\/share.php?url=https:\/\/vc.ru\/services\/190625-rossiyskiy-startap-v-oblasti-issledovaniy-dnk-genotek-privlek-4-mln-ot-yandeksa-i-drugih-investorov&title=\u0420\u043e\u0441\u0441\u0438\u0439\u0441\u043a\u0438\u0439 \u0441\u0442\u0430\u0440\u0442\u0430\u043f \u0432 \u043e\u0431\u043b\u0430\u0441\u0442\u0438 \u0438\u0441\u0441\u043b\u0435\u0434\u043e\u0432\u0430\u043d\u0438\u0439 \u0414\u041d\u041a Genotek \u043f\u0440\u0438\u0432\u043b\u0451\u043a $4\u00a0\u043c\u043b\u043d \u043e\u0442 \u00ab\u042f\u043d\u0434\u0435\u043a\u0441\u0430\u00bb \u0438 \u0434\u0440\u0443\u0433\u0438\u0445 \u0438\u043d\u0432\u0435\u0441\u0442\u043e\u0440\u043e\u0432″,»short_name»:»VK»,»title»:»\u0412\u041a\u043e\u043d\u0442\u0430\u043a\u0442\u0435″,»width»:600,»height»:450},»twitter»:{«url»:»https:\/\/twitter. com\/intent\/tweet?url=https:\/\/vc.ru\/services\/190625-rossiyskiy-startap-v-oblasti-issledovaniy-dnk-genotek-privlek-4-mln-ot-yandeksa-i-drugih-investorov&text=\u0420\u043e\u0441\u0441\u0438\u0439\u0441\u043a\u0438\u0439 \u0441\u0442\u0430\u0440\u0442\u0430\u043f \u0432 \u043e\u0431\u043b\u0430\u0441\u0442\u0438 \u0438\u0441\u0441\u043b\u0435\u0434\u043e\u0432\u0430\u043d\u0438\u0439 \u0414\u041d\u041a Genotek \u043f\u0440\u0438\u0432\u043b\u0451\u043a $4\u00a0\u043c\u043b\u043d \u043e\u0442 \u00ab\u042f\u043d\u0434\u0435\u043a\u0441\u0430\u00bb \u0438 \u0434\u0440\u0443\u0433\u0438\u0445 \u0438\u043d\u0432\u0435\u0441\u0442\u043e\u0440\u043e\u0432″,»short_name»:»TW»,»title»:»Twitter»,»width»:600,»height»:450},»telegram»:{«url»:»tg:\/\/msg_url?url=https:\/\/vc.ru\/services\/190625-rossiyskiy-startap-v-oblasti-issledovaniy-dnk-genotek-privlek-4-mln-ot-yandeksa-i-drugih-investorov&text=\u0420\u043e\u0441\u0441\u0438\u0439\u0441\u043a\u0438\u0439 \u0441\u0442\u0430\u0440\u0442\u0430\u043f \u0432 \u043e\u0431\u043b\u0430\u0441\u0442\u0438 \u0438\u0441\u0441\u043b\u0435\u0434\u043e\u0432\u0430\u043d\u0438\u0439 \u0414\u041d\u041a Genotek \u043f\u0440\u0438\u0432\u043b\u0451\u043a $4\u00a0\u043c\u043b\u043d \u043e\u0442 \u00ab\u042f\u043d\u0434\u0435\u043a\u0441\u0430\u00bb \u0438 \u0434\u0440\u0443\u0433\u0438\u0445 \u0438\u043d\u0432\u0435\u0441\u0442\u043e\u0440\u043e\u0432″,»short_name»:»TG»,»title»:»Telegram»,»width»:600,»height»:450},»odnoklassniki»:{«url»:»http:\/\/connect.
com\/intent\/tweet?url=https:\/\/vc.ru\/services\/190625-rossiyskiy-startap-v-oblasti-issledovaniy-dnk-genotek-privlek-4-mln-ot-yandeksa-i-drugih-investorov&text=\u0420\u043e\u0441\u0441\u0438\u0439\u0441\u043a\u0438\u0439 \u0441\u0442\u0430\u0440\u0442\u0430\u043f \u0432 \u043e\u0431\u043b\u0430\u0441\u0442\u0438 \u0438\u0441\u0441\u043b\u0435\u0434\u043e\u0432\u0430\u043d\u0438\u0439 \u0414\u041d\u041a Genotek \u043f\u0440\u0438\u0432\u043b\u0451\u043a $4\u00a0\u043c\u043b\u043d \u043e\u0442 \u00ab\u042f\u043d\u0434\u0435\u043a\u0441\u0430\u00bb \u0438 \u0434\u0440\u0443\u0433\u0438\u0445 \u0438\u043d\u0432\u0435\u0441\u0442\u043e\u0440\u043e\u0432″,»short_name»:»TW»,»title»:»Twitter»,»width»:600,»height»:450},»telegram»:{«url»:»tg:\/\/msg_url?url=https:\/\/vc.ru\/services\/190625-rossiyskiy-startap-v-oblasti-issledovaniy-dnk-genotek-privlek-4-mln-ot-yandeksa-i-drugih-investorov&text=\u0420\u043e\u0441\u0441\u0438\u0439\u0441\u043a\u0438\u0439 \u0441\u0442\u0430\u0440\u0442\u0430\u043f \u0432 \u043e\u0431\u043b\u0430\u0441\u0442\u0438 \u0438\u0441\u0441\u043b\u0435\u0434\u043e\u0432\u0430\u043d\u0438\u0439 \u0414\u041d\u041a Genotek \u043f\u0440\u0438\u0432\u043b\u0451\u043a $4\u00a0\u043c\u043b\u043d \u043e\u0442 \u00ab\u042f\u043d\u0434\u0435\u043a\u0441\u0430\u00bb \u0438 \u0434\u0440\u0443\u0433\u0438\u0445 \u0438\u043d\u0432\u0435\u0441\u0442\u043e\u0440\u043e\u0432″,»short_name»:»TG»,»title»:»Telegram»,»width»:600,»height»:450},»odnoklassniki»:{«url»:»http:\/\/connect. ok.ru\/dk?st.cmd=WidgetSharePreview&service=odnoklassniki&st.shareUrl=https:\/\/vc.ru\/services\/190625-rossiyskiy-startap-v-oblasti-issledovaniy-dnk-genotek-privlek-4-mln-ot-yandeksa-i-drugih-investorov»,»short_name»:»OK»,»title»:»\u041e\u0434\u043d\u043e\u043a\u043b\u0430\u0441\u0441\u043d\u0438\u043a\u0438″,»width»:600,»height»:450},»email»:{«url»:»mailto:?subject=\u0420\u043e\u0441\u0441\u0438\u0439\u0441\u043a\u0438\u0439 \u0441\u0442\u0430\u0440\u0442\u0430\u043f \u0432 \u043e\u0431\u043b\u0430\u0441\u0442\u0438 \u0438\u0441\u0441\u043b\u0435\u0434\u043e\u0432\u0430\u043d\u0438\u0439 \u0414\u041d\u041a Genotek \u043f\u0440\u0438\u0432\u043b\u0451\u043a $4\u00a0\u043c\u043b\u043d \u043e\u0442 \u00ab\u042f\u043d\u0434\u0435\u043a\u0441\u0430\u00bb \u0438 \u0434\u0440\u0443\u0433\u0438\u0445 \u0438\u043d\u0432\u0435\u0441\u0442\u043e\u0440\u043e\u0432&body=https:\/\/vc.ru\/services\/190625-rossiyskiy-startap-v-oblasti-issledovaniy-dnk-genotek-privlek-4-mln-ot-yandeksa-i-drugih-investorov»,»short_name»:»Email»,»title»:»\u041e\u0442\u043f\u0440\u0430\u0432\u0438\u0442\u044c \u043d\u0430 \u043f\u043e\u0447\u0442\u0443″,»width»:600,»height»:450}},»isFavorited»:false}
ok.ru\/dk?st.cmd=WidgetSharePreview&service=odnoklassniki&st.shareUrl=https:\/\/vc.ru\/services\/190625-rossiyskiy-startap-v-oblasti-issledovaniy-dnk-genotek-privlek-4-mln-ot-yandeksa-i-drugih-investorov»,»short_name»:»OK»,»title»:»\u041e\u0434\u043d\u043e\u043a\u043b\u0430\u0441\u0441\u043d\u0438\u043a\u0438″,»width»:600,»height»:450},»email»:{«url»:»mailto:?subject=\u0420\u043e\u0441\u0441\u0438\u0439\u0441\u043a\u0438\u0439 \u0441\u0442\u0430\u0440\u0442\u0430\u043f \u0432 \u043e\u0431\u043b\u0430\u0441\u0442\u0438 \u0438\u0441\u0441\u043b\u0435\u0434\u043e\u0432\u0430\u043d\u0438\u0439 \u0414\u041d\u041a Genotek \u043f\u0440\u0438\u0432\u043b\u0451\u043a $4\u00a0\u043c\u043b\u043d \u043e\u0442 \u00ab\u042f\u043d\u0434\u0435\u043a\u0441\u0430\u00bb \u0438 \u0434\u0440\u0443\u0433\u0438\u0445 \u0438\u043d\u0432\u0435\u0441\u0442\u043e\u0440\u043e\u0432&body=https:\/\/vc.ru\/services\/190625-rossiyskiy-startap-v-oblasti-issledovaniy-dnk-genotek-privlek-4-mln-ot-yandeksa-i-drugih-investorov»,»short_name»:»Email»,»title»:»\u041e\u0442\u043f\u0440\u0430\u0432\u0438\u0442\u044c \u043d\u0430 \u043f\u043e\u0447\u0442\u0443″,»width»:600,»height»:450}},»isFavorited»:false}
3979 просмотров
| МИНИСТР ОБОРОНЫ РОССИЙСКОЙ ФЕДЕРАЦИИ | ||
| Генерал армии Шойгу Сергей Кужугетович | ||
| ЗАМЕСТИТЕЛИ МИНИСТРА ОБОРОНЫ РОССИИ | ||
| Начальник Генерального штаба Вооруженных Сил РФ — первый заместитель Министра обороны Российской Федерации генерал армии Герасимов Валерий Васильевич |
||
| Первый заместитель Министра обороны Российской Федерации Цаликов Руслан Хаджисмелович |
||
| Статс-секретарь — заместитель Министра обороны Российской Федерации Панков Николай Александрович |
||
| Заместитель Министра обороны Россйской Федерации — начальник Главного военно-политического управления Вооруженных Сил Российской Федерации генерал-полковник Картаполов Андрей Валериевич | ||
| Заместитель Министра обороны Российской Федерации — руководитель Аппарата Министра обороны Российской Федерации генерал-полковник Садовенко Юрий Эдуардович |
||
| Заместитель Министра обороны Российской Федерации генерал армии Булгаков Дмитрий Витальевич |
||
| Заместитель Министра обороны Российской Федерации генерал-лейтенант Евкуров Юнус-Бек Баматгиреевич |
||
| Заместитель Министра обороны Российской Федерации Иванов Тимур Вадимович |
||
| Заместитель Министра обороны Российской Федерации Криворучко Алексей Юрьевич |
||
| Заместитель Министра обороны Российской Федерации генерал армии Попов Павел Анатольевич |
||
| Заместитель Министра обороны Российской Федерации Фомин Александр Васильевич |
||
| Заместитель Министра обороны Российской Федерации Шевцова Татьяна Викторовна |
||
ГЕНЕРАЛЬНЫЙ ШТАБ ВООРУЖЕННЫХ СИЛ РОССИЙСКОЙ ФЕДЕРАЦИИ |
||
| Главное оперативное управление ГШ ВС РФ | ||
| Главное управление ГШ ВС РФ | ||
| Главное организационно-мобилизационное управление ГШ ВС РФ | ||
| Главное управление связи ВС РФ | ||
| Национальный центр управления обороной Российской Федерации | ||
| Управление начальника войск радиоэлектронной борьбы ВС РФ | ||
| Военно-топографическое управление ГШ ВС РФ | ||
| Восьмое управление ГШ ВС РФ | ||
| Управление оперативной подготовки ВС РФ | ||
| Управление (строительства и развития системы применения беспилотных летательных аппаратов) ГШ ВС РФ |
||
| Архивная служба ВС РФ | ||
| ГЛАВНЫЕ УПРАВЛЕНИЯ | ||
| Главное управление боевой подготовки ВС РФ | ||
| Главное управление военной полиции Минобороны России | ||
| Главное управление кадров Минобороны России | ||
| Главное управление международного военного сотрудничества Минобороны России |
||
| Главное управление вооружения ВС РФ | ||
| Главное автобронетанковое управление Минобороны России | ||
| Главное ракетно-артиллерийское управление Минобороны России | ||
| Главное управление начальника Железнодорожных войск Минобороны России | ||
| Главное военно-медицинское управление Минобороны России | ||
| Главное управление научно-исследовательской деятельности и технологического сопровождения передовых технологий (инновационных исследований) Минобороны России |
||
| Главное управление контрольной и надзорной деятельности Министерства обороны Российской Федерации | ||
| Главное военно-политическое управление Вооруженных Сил Российской Федерации | ||
| УПРАВЛЕНИЯ | ||
| Управление начальника войск радиационной, химической и биологической защиты ВС РФ | ||
| Управление начальника инженерных войск ВС РФ | ||
| Управление службы войск и безопасности военной службы Минобороны России |
||
| Управление государственного надзора за ядерной и радиационной безопасностью Минобороны России | ||
| Управление физической подготовки и спорта ВС РФ | ||
| Управление военных представительств Минобороны России | ||
| Управление Минобороны России по увековечению памяти погибших при защите Отечества |
||
| Управление Минобороны России по контролю за выполнением договоров (Национальный центр по уменьшению ядерной опасности) | ||
| Управление метрологии ВС РФ | ||
| Федеральное управление накопительно-ипотечной системы жилищного обеспечения военнослужащих |
||
| Управление делами Минобороны России | ||
| Организационное управление Минобороны России | ||
| Управление Минобороны России по работе с обращениями граждан (общественная приёмная Министра обороны Российской Федерации) |
||
| Контрольное управление Минобороны России | ||
| Управление интеллектуальной собственности, военно-технического сотрудничества и экспертизы поставок вооружения и военной техники Минобороны России |
||
| Управление перспективных межвидовых исследований и специальных проектов Минобороны России | ||
| Управление заказов по совершенствованию технической основы системы управления ВС РФ | ||
| Государственная экспертиза Минобороны России | ||
| Военно-научный комитет ВС РФ | ||
| Научно-технический комитет (развития вооружений) | ||
| ДЕПАРТАМЕНТЫ | ||
| Штаб материально-технического обеспечения Вооруженных Сил Российской Федерации | ||
| Департамент ресурсного обеспечения Минобороны России | ||
| Департамент транспортного обеспечения Минобороны России | ||
| Департамент планирования и координации обустройства войск (сил) Минобороны России | ||
| Департамент строительства Минобороны России | ||
| Департамент жилищного обеспечения и управления жилищным фондом Минобороны России | ||
| Департамент военного имущества Минобороны России | ||
| Департамент финансового обеспечения Минобороны России | ||
| Департамент финансового планирования Министерства обороны Российской Федерации | ||
| Департамент социальных гарантий Министерства обороны Российской Федерации | ||
| Департамент эксплуатационного содержания и обеспечения коммунальными услугами воинских частей и организаций Минобороны России | ||
| Департамент информации и массовых коммуникаций Минобороны России | ||
| Пресс-секретарь Министра обороны Российской Федерации | ||
| Правовой департамент Минобороны России | ||
| Протокольно-координационный департамент Министерства обороны Российской Федерации | ||
| Департамент государственных закупок Минобороны России | ||
| Департамент Министерства обороны Российской Федерации по обеспечению государственного оборонного заказа | ||
| Департамент военно-экономического анализа Министерства обороны Российской Федерации | ||
| Департамент аудита государственных контрактов Министерства обороны Российской Федерации | ||
| Департамент ведомственного финансового контроля и аудита Министерства обороны Российской Федерации | ||
| Департамент финансового мониторинга государственного оборонного заказа Минобороны России | ||
| Департамент культуры Минобороны России | ||
| Департамент информационных систем Министерства обороны Российской Федерации | ||
| Департамент психологической работы Министерства обороны Российской Федерации | ||
| СЛУЖБЫ | ||
| Служба безопасности полетов авиации ВС РФ | ||
| Военно-оркестровая служба ВС РФ | ||
| Военно-геральдическая служба ВС РФ | ||
| Гидрометеорологическая служба ВС РФ | ||
| ГЛАВНЫЕ КОМАНДОВАНИЯ | ||
| Главное командование Сухопутных войск | ||
| Главное командование Воздушно-космических сил | ||
| Главное командование Военно-Морского Флота | ||
| КОМАНДОВАНИЯ | ||
| Командование Ракетных войск стратегического назначения | ||
| Командование Воздушно-десантных войск | ||
Повествовательный обзор и метаанализ
напоминают действие b-лактамных антибиотиков. PLoS One 2014;
PLoS One 2014;
9 (9): e106967.
[27] Майтан Дж., Бохова Дж., Хорняцкова М., Клаудины Дж., Майтан В.
Антибиотикопленочные эффекты меда против раневых патогенов
Proteus mirabilis и Enterobacter cloacae. Phytother Res
2014; 28 (1): 69e75.
[28] Лу Дж., Тернбулл Л., Берк С.М., Лю М., Картер Д.А.,
Шлотхауэр Р.С. и др. Мед типа манука может уничтожить
биопленок, продуцируемых штаммами Staphylococcus aureus с
различными способностями к формированию биопленок.PeerJ 2014; 2: e326.
[29] Ансари М.Дж., Аль-Гамди А., Усмани С., Аль-Вайли Н.С., Шарма Д.,
Нуру А. и др. Влияние меда из мармелада на рост Candida albicans
и формирование биопленки. Arch Med Res 2013; 44 (5):
352e60.
[30] Мерколл П., Йонассен Т., Вад М.Э., Жанссон С.Л., Мелби К.К.
Бактерии, биопленка и мед: исследование воздействия меда
на «планктонные» и встроенные в биопленку хронические
раневые бактерии. Scand J Infect Dis 2009; 41 (5): 341e7.
Scand J Infect Dis 2009; 41 (5): 341e7.
[31] Атротт Дж., Хенле Т. Метилглиоксаль в меде Манука ecor-
имеет антибактериальные свойства. Чешский J Food Sci
2009; 27: S163e5.
[32] Стивенс Дж. М., Шлотхауэр Р. К., Моррис Б. Д., Ян Д.,
Фернли Л., Гринвуд Д. Р. и др. Фенольные соединения и
метилглиоксаль в некоторых новозеландских медах манука и канука
. Food Chem 2010; 120 (1): 78e86.
[33] Майтан Дж. Метилглиоксаль — потенциальный фактор риска меда манука
при заживлении диабетических язв.Evid Based Comple-
, Альтернативная медицина 2011; 2011: 295494.
[34] Катираи Ф., Махмоди Р., Мардани К., Бабаи Э. Противогрибковое средство
активность иранских медоносных пчел против Candida, Aspergillus
видов и Trichophyton Rubrum. J Food Process Preserv
2014; 38 (5): 2078e82.
[35] Канонико Б., Кандираччи М., Читтерио Б., Курчи Р., Скварцони С.,
Маццони А. и др. Медофлавоноиды подавляют морфогенез Candida albicans
, влияя на поведение ДНК и дериальную функцию митохонов. Future Microbiol 2014; 9 (4): 445e56.
[36] Захур М., Наз С., Сангин М. Антибактериальная, противогрибковая и
антиоксидантная активность меда, собранного из Тимергара
(Дир, Пакистан). Pak J Pharm Sci 2014; 27 (1): 45e50.
[37] Brudzynski K, Abubaker K, St-Martin L, Castle A. Re-
исследует роль перекиси водорода в бактериостатической
и бактерицидной активности меда. Front Microbiol 2011;
2: 213.
[38] Найт А.Лечебные эффекты меда. Плимут
Student Sci 2013; 6 (1): 376e85.
[39] Мандал М.Д., Мандал С. Мед: его лечебные свойства и
антибактериальная активность. Азиатский Pac J Trop Biomed 2011; 1 (2):
154e60.
[40] Банг Л.М., Банттинг С., Молан П. Влияние разбавления на скорость
производства перекиси водорода в меде и его
последствия для заживления ран. J. Альтернативное дополнение Med
2003; 9 (2): 267e73.
[41] Kwakman PH, de Boer L, Ruyter-Spira CP, Creemers-
Molenaar T. , Helsper JP, Vandenbroucke-Grauls CM, et al.
, Helsper JP, Vandenbroucke-Grauls CM, et al.
Мед медицинского назначения, обогащенный антимикробными пептидами
, обладает повышенной активностью в отношении устойчивых к антибиотикам патогенов.
. Eur J Clin Microbiol Infect Dis 2011; 30 (2): 251e7.
[42] Банеян-Боружени С., Мобини Г.Р., Пургейсари Б., Валиди М.
Сравнение действия меда и миконазола против
Candida albicans in vitro. Adv Biomed Res 2013; 30 (2): 57.
[43] Уайт Дж. У., Суберс М. Х., Шепарц А. И.. Идентификация ингибина
, антибактериального фактора меда, как пероксида водорода
и его происхождения в системе глюкозеоксидазы меда.
Biochim Biophys Acta 1963; 73: 57e70.
[44] Schepartz AI, Subers MH. Глюкозооксидаза меда: I.
Очистка и некоторые общие свойства фермента.
Biochim Biophys Acta 1964; 85: 228e37.
[45] Брудзинский К. Влияние перекиси водорода на антибактериальную
активность канадского меда. Can J Microbiol 2006; 52 (12):
1228e37.
[46] Чен С., Кэмпбелл Л.Т., Блэр С.Е., Картер Д.А. Влияние
стандартных процедур термофильтрации на антимикробную активность
и уровни перекиси водорода в меде
.Front Microbiol 2012; 3: 265.
[47] Майтан Дж., Бохова Дж., Прочазка Е., Клаудины Дж. Метилглиоксаль
может влиять на накопление перекиси водорода в меде манука
за счет ингибирования глюкозооксидазы. J Med
Food 2014; 17 (2): 290e3.
[48] Айриш Дж., Блэр С., Картер Д.А. Антибактериальная активность
меда, полученного из австралийской флоры. PLoS One 2011; 6 (3):
e18229.
[49] Kretavi
cius J, Kurtinaitien _
eB, Ra
cys J,
Ceksteryt _
eV.
Инактивация глюкозооксидазы при термообработке
кристаллизация меда.
Земдирбыст _
e Agric 2010; 97 (4):
115e22.
[50] Майтан Дж., Клаудины Дж., Бохова Дж., Кохутова Л., Дзурова М. ,
,
Седива М. и др. Метилглиоксаль-индуцированные модификации значительных белковых компонентов пчел
в меде манука
: возможные терапевтические последствия. Фитотерапия
2012; 83 (4): 671e7.
[51] Molan PC. Антибактериальная активность меда. 2. Вариация
по антибактериальной активности. Пчелиный мир
1992; 73 (2): 59e76.
[52] Kwakman PH, Te Velde AA, de Boer L, Vandenbroucke-
Grauls CM, Zaat SA. Два основных лекарственных меда обладают
различных механизмов бактерицидной активности. PLoS One
2011; 6 (3): e17709.
[53] Бучекова М., Валачева И., Кохутова Л., Прохазка Е.,
Клаудины Ю., Майтан Дж.Глюкоза оксидазы медоносной пчелы выражается в
у рабочих пчел и сравнительный анализ ее содержания
и H
2
O
2
-опосредованная антибактериальная активность в
натуральных медах. Naturwissenschaften 2014; 101 (8): 661e70.
[54] Molan PC. Возможности меда при лечении ран
Возможности меда при лечении ран
и ожогов. Am J Clin Dermatol 2001; 2 (1): 13e9.
[55] Росситер К., Купер А.Дж., Фогели Д., Лвалид Б.А. Мед
способствует ангиогенной активности в анализе кольца аорты крысы.J
Уход за ранами 2010; 19 (10): 440,442e440,446.
[56] Селл С.А., Вулф П.С., Спенс А.Дж., Родригес И.А., МакКул Дж.М.,
Петрелла Р.Л. и др. Предварительное исследование потенциала
меда манука и богатой тромбоцитами плазмы при заживлении ран.
Int J Biomater 2012; 2012: 313781.
[57] Аль-Вайли Н., Салом К., Аль-Гамди А.А. Мед для заживления ран, язв и ожогов
; данные, подтверждающие его использование в клинической практике
. Научный журнал WorldJournal 2011; 11: 766e87.
[58] Сингх М.П., Чурасия Х.Р., Агарвал М., Малхотра А., Шарма М.,
Шарма Д. и др. Мед как дополнительное лекарство: обзор
. IJPBS 2012; 3 (2): 12e31.
[59] Аггад Х., Гемур Д. Антибактериальная активность меда. Med
Aromat Plants 2014; 3: 2.
[60] Maddocks SE, Jenkins RE. Мед: сладкое решение растущей проблемы устойчивости к противомикробным препаратам
? Будущее
Microbiol 2013; 8 (11): 1419e29.
[61] Джордж Н.М., Каттинг К.Ф.Антибактериальный мед (Medihoney):
активность in vitro против клинических изолятов MRSA, VRE и
других мультирезистентных грамотрицательных организмов, включая
Pseudomonas aeruginosa. Раны 2007; 19 (9): 231e6.
[62] Стюарт Дж. А., МакГрейн О. Л., Уэдмор И. С.. Уход за ранами в пустыне
: есть ли доказательства наличия меда? Wilderness Envi-
ron Med 2014; 25 (1): 103e10.
[63] Алам Ф, Ислам М.А., Ган Ш., Халил М.И. Мед: потенциальное терапевтическое средство
для лечения диабетических ран.Evid
на основе Complement Altern Med 2014; 2014: 169130.
[64] Карайил С., Дешпанде С.Д., Коппикар Г.В. Влияние меда на
организмов с множественной лекарственной устойчивостью и его синергетическое действие
18 A. Oryan et al.
Цитируйте эту статью в прессе как: Oryan A et al., Биологические свойства и терапевтическое действие меда при заживлении ран: повествовательный обзор и метаанализ
, Journal of Tissue Viability (2016), http: // dx .doi.org / 10.1016 / j.jtv.2015.12.002
Границы | Решения шмелей и медоносных пчел о кормлении на индивидуальном уровне и на уровне колонии в связи с балансированием потребностей в питательных веществах
Введение
Что поесть? Где собирать корм и как долго? Принятие решений в контексте поведения, связанного с поиском пищи, может быть сложным процессом, включающим несколько сенсорных модальностей, определенные формы обучения и памяти, а также интеграцию множества различных источников внешней окружающей среды и внутренней физиологической информации.Теория оптимального кормодобывания (OFT) делает прогнозы относительно типов решений, которые принимают отдельные животные, чтобы минимизировать затраты энергии и максимизировать потребление собственных питательных веществ, некоторые из которых были подтверждены исследованиями на различных таксонах животных (Pyke, 1984; Sih and Кристенсен, 2001). Тем не менее, OFT подвергается резкой критике за игнорирование особенностей жизненного цикла, потому что добыча пищи может происходить в контекстах, выходящих за рамки удовлетворения насущных потребностей людей, например, родительская забота и накопление пищи (Pierce and Ollason, 1987).
Решения о собирательстве требуют еще большего уровня сложности при рассмотрении социальных животных, особенно высоко социальных видов, для которых важными факторами являются инклюзивная приспособленность и групповые выгоды (Galef and Giraldeau, 2001; Kay, 2002). Эусоциальные насекомые с репродуктивным разделением труда, которое включает в себя каста непроизводящих рабочих, такую как муравьи и медоносные пчелы, представляют собой крайний случай, когда собиратели часто очень специализированы на поведении при сборе пищи (Wilson, 1971).Однако этот сбор продуктов питания часто не зависит от их собственных индивидуальных потребностей в питании, и собранные продукты часто не потребляются напрямую, а вместо этого используются для удовлетворения потребностей колонии в питании. Ярким примером является медоносный муравей Myrmecocystus , у которого особая каста особей служит живыми приемниками пищи (Burgett and Young, 1974), накапливая огромное количество секретов сладких растений в своих культурах, но не потребляя и не переваривая подавляющее большинство. этой пищи для удовлетворения собственных физиологических потребностей.
Эусоциальные насекомые произошли от одиночных видов, у которых особи должны были реагировать на свои собственные внутренние физиологические сигналы, чтобы прокормиться, чтобы подпитывать личные репродуктивные возможности (Ament et al., 2010). В ходе эволюции социальности также должен был происходить переход от принятия решений о добыче пищи, основанных на индивидуальных потребностях, к решениям, основанным на потребностях социальной группы (Behmer, 2009b; Boomsma and Gawne, 2018). Из всего разнообразия современных видов насекомых, встречающихся сегодня, есть виды с высокой эусоциальностью, у которых колонии очень большие, а отдельные рабочие редко воспроизводятся, а также примитивно эусоциальные виды, у которых колонии меньше, а отдельные рабочие сохраняют некоторую репродуктивную способность (Wilson, 1971).Таким образом, возможно, удастся выявить различия в том, как социальные насекомые с разными формами социальности уравновешивают индивидуальные решения относительно кормодобывания на уровне колонии. Если энергетический баланс колонии нарушается, происходит ли противодействующая реакция на уровне группы (изменение разделения труда) или на индивидуальном уровне (изменение индивидуального выбора корма)? И отличаются ли эти ответы у видов с разным уровнем социальности? Мы стремились восполнить этот пробел в знаниях, изучая поведенческие реакции двух видов пчел с разными формами социальной организации.
В пределах хорошо изученного пчелиного подсемейства Apinae, Apis mellifera медоносные пчелы являются классическим примером высокоэусоциального вида со специализированными работниками (Johnson, 2010), в то время как у шмелей Bombus terrestris есть небольшие семьи с более гибким поведением рабочие (Crall et al., 2018). A. mellifera живут в многолетних колониях, запасающих пищу, состоящих из десятков тысяч рабочих женщин, большинство из которых никогда не откладывают яйца в течение нормального цикла колоний (Winston, 1991).Медоносные пчелы, собирающие медоносные пчелы, как известно, в высшей степени специализируются на сборе пыльцы, нектара или непищевых веществ, таких как вода или прополис (Page and Fondrk, 1995). Напротив, колонии B. terrestris являются однолетними, с небольшими запасами пыльцы и нектара, и по мере уменьшения количества колоний осенью рабочие вступают в борьбу за доминирование, и некоторые из них откладывают неоплодотворенные яйца (Goulson, 2010). Хотя более крупные рабочие обычно добывают корм, а более мелкие — заботятся о расплодах, отдельные сборщики не имеют такой узкой специализации, как медоносные пчелы (Goulson et al., 2002; Crall et al., 2018). Кроме того, у шмелей есть только относительно простая система общения во время кормодобывания по сравнению с изощренным танцевальным языком медоносных пчел (Dornhaus and Chittka, 2001). Принятие решений на уровне колонии, например, выделение колонии собирателей для пыльцы или нектара, также широко изучалось как у медоносных пчел, так и у шмелей (Cartar, 1992; Seeley, 1995; Hagbery and Nieh, 2012; Leonhardt and Blüthgen, 2012). ; Konzmann, Lunau, 2014; Vaudo et al., 2014; Ruedenauer et al., 2015). Принятие индивидуальных решений во время походов за кормом также широко изучается на медоносных пчелах и шмелях, обеспечивая важные проверки прогнозов OFT (Heinrich, 1983; Hodges, 1985; Wells and Wells, 1986; Lihoreau et al., 2012; Katz and Naug, 2015). Однако, насколько нам известно, в нескольких исследованиях одновременно рассматривались ответы как на индивидуальном уровне, так и на уровне колонии (Schulz et al., 1998).
В качестве руководящей основы для этого исследования мы использовали геометрическую схему для сбалансированного питания белков (P) и углеводов (C), разработанную на основе исследований на одиночных насекомых (Lee et al., 2002; Бехмер, 2009а; Raubenheimer et al., 2009). Геометрическая схема предсказывает, что выбор продуктов питания для ключевых макроэлементов, таких как P и C, будет геометрически масштабироваться в оптимальном соотношении, которое поддерживает индивидуальную репродуктивную пригодность. Геометрическая структура также может быть распространена на целые колонии (Behmer, 2009b; Dussutour and Simpson, 2009; Lihoreau et al., 2014). Это исследование расширяет результаты исследования баланса фосфора и углерода в изолированных когортах медоносных пчел, содержащихся в клетках (Altaye et al., 2010; Pirk et al., 2010; Paoli et al., 2014а, б) и шмелей (Stabler et al., 2015) на весь уровень колонии.
Мы изменили входы фосфора и углерода в колонии обоих видов пчел и изучили, как это повлияло на (1) распределение колоний фуражиров по станциям кормления с предвзятым отношением фосфора и углерода и (2) индивидуальные предпочтения (на основании выбора и загрузки корма в разных условиях). станции питания) с учетом диапазона концентраций P и C. Мы предположили, что и медоносные пчелы, и шмели будут регулировать разделение труда в колониях и выделять больше собирателей для P и / или C в зависимости от потребностей колонии.Было предсказано, что индивидуальные предпочтения будут смещены в сторону более высоких концентраций P и C для собирателей из колоний, более серьезно лишенных P или C, соответственно, для удовлетворения краткосрочных потребностей колонии, а не для долгосрочного оптимального кормления. Мы также предсказали, что из-за различий в уровне социальной жизни и запасах пищи шмели будут демонстрировать более сильную индивидуальную реакцию кормления на баланс питательных веществ, тогда как отдельные медоносные пчелы будут более фиксированными в ответ на изменения питательных веществ в колониях.
Материалы и методы
Подготовка колонии медоносных пчел
Двадцать четыре колонии медоносных пчел ( A. mellifera ligustica ) были стандартизированы в ульях-ядрах с ~ 5000 рабочих, гнездо для расплода в трех центральных рамках и хранилища меда в двух боковых рамках (т.е. всего 5 рамок). Этот размер колонии представляет собой фазу активного роста в жизненном цикле колонии (т. Е. Не старение или размножение) (Johnson, 2010). После стандартизации (9 октября 2014 г.) все колонии были перемещены на 3 недели в место 4.На расстоянии 2 км, чтобы пчелы не возвращались в места бывшего улья, и чтобы восстановить баланс между рабочими, выводком и магазинами.
За день до экспериментов с медоносными пчелами каждой колонии случайным образом назначали одну из трех обработок пыльцы: дефицит пыльцы (P–), контроль пыльцы (P) или избыток пыльцы (P +). Восемь колоний были помещены в пустую соту в середине гнезда расплода, в то время как любые пыльцевые клетки в этих колониях были изолированы с помощью расплавленного воска (обработка P–). Запечатывание пыльцы подобно естественному запечатыванию медоносных пчел, которое называется укупоркой или погребением (vanEngelsdorp et al., 2009). Восемь колоний были контрольными, с которыми не проводили никаких манипуляций (обработка P). В восьми других колониях манипулировали путем введения соты, полностью заполненной пыльцой, в середину гнезда для расплода (обработка P +). Эти соты для пыльцы были сделаны путем заполнения предварительно взвешенных пустых вытянутых сот смешанными гранулами пыльцы. Гранулы прессовали в клетки рабочими пчелами в большой колонии в течение 1 дня. Впоследствии каждую гребенку повторно взвешивали (в среднем 581 г добавленных гранул ± 29,7 SE; n = 8) и хранили в холодильнике до использования.
Чтобы ввести в гнездо расплода пустую соту (П–) или полную соту (П +), вынули одну из боковых сот с медом. Удаление меда рассматривалось как манипуляция уровнем углеводного питания (лечение C–). Таким образом, дизайн позволил оценить манипуляции в улье как с P (уровни: меньше, нормальный и больше), так и с C (уровни: меньше и нормально) хранилищами (Таблица 1).
Таблица 1 . Экспериментальные манипуляции применительно к колониям.
Подготовка колонии шмелей
Двадцать четыре колонии шмелей европейских желтохвостых шмелей ( B.terrestris) были получены от коммерческого селекционера (Biobee, Sde Eliyahu, Израиль). Они были приготовлены 1 декабря и доставлены 3 декабря 2014 года. Такой размер колонии представляет собой фазу активного роста в жизненном цикле колонии, поскольку еще не выращивали трутней или гинекологов. После эксперимента колонии были убиты, чтобы свести к минимуму вероятность распространения патогенов в местных популяциях пчел (Aizen et al., 2018). Последующие показатели были собраны по рабочим колонии, расплоду и хранению корма (пыльца и нектар) (Приложение 1).
Стандартные кормушки для сахарозы в улье (2 л; 50% раствор сахарозы) и пирожки с пыльцой в улье (50 г; 90% пыльцы с 10% раствором сахарозы) были извлечены из восьми ящиков для колоний шмелей. Эти колонии были базовыми контрольными колониями, которым необходимо было кормить как белком (P; пыльцевый рацион), так и углеводами (C; раствор сахарозы) в соответствии с нормальной потребностью (соотношение P: C). Восемь дополнительных колоний оставили с поставляемыми в улье пирожками с пыльцой (P +), но не с кормушками для сахарозы (C). Еще восемь колоний сохранили в улье питатели сахарозы (C +), но не пирожки с пыльцой (P).Таким образом, дизайн позволял манипулировать колониями как диеты P (уровни: нормальные и более), так и диеты C (уровни: нормальные и более) (Таблица 1).
Различия в манипулировании колониями медоносных пчел и шмелей
Поскольку в эксперименте участвовали два разных таксона пчел с врожденными различиями в характеристиках хранения пыльцы и нектара, точно параллельные манипуляции с колониями были невозможны. Семейства медоносных пчел хранят многодневный буфер с пыльцой в сотах, чтобы обезопасить кормление пчел.Пчелы-медсестры переваривают пыльцу, чтобы получить желе для кормления личинок (Wright et al., 2018). Напротив, шмели собирают пыльцу для быстрого обеспечения клеток расплода, поскольку их личинки напрямую поедают пыльцу (Stabler et al., 2015). Это несоответствие отражено экспериментальными манипуляциями, чтобы включить уровни обработки меньшего, нормального и большего количества P в улье в качестве лечения медоносных пчел, в то время как шмели включали только уровни обработки нормальный и более высокий уровень P в улье, потому что удаление пыльцы из колоний шмелей было неосуществимо.Во-вторых, колонии медоносных пчел хранят большое количество меда, в то время как шмели содержат относительно небольшое количество медовых горшков. По сравнению с сотами для пчел, горшками с медом для шмелей манипулировать сложнее. Таким образом, запасами углеводов в колониях медоносных пчел манипулировали, удаляя соты (C–), но в семьях шмелей, добавляя сахарозную кормушку (C +), в то время как исходное состояние колонии (C) присутствовало для обоих видов (Таблица 1) . Обработку колоний можно резюмировать как P + C–, PC и P – C– для медоносных пчел и P + C, PC и PC + для шмелей.
Экспериментальный сайт и установка
Эксперименты проводились на экспериментальной ферме имени Джозефа Маргулеаса факультета сельского хозяйства, продовольствия и окружающей среды Еврейского университета в Иерусалиме в Реховоте (31 ° 54′16,02 ″ с.ш., 34 ° 47′50,09 ″ в.д.). Использовали 24 туннельных клетки размером 2 × 3 × 2 м (длина × ширина × высота) с сетчатым покрытием с ячейками 1,3 мм. Каждая колония имела доступ к ведру с водой с пробковыми поплавками и ареной для кормления, состоящей из двух платформ, покрытых листами белой бумаги размером 50 × 50 см, для подачи каждой колонии пятью пыльцами и пятью кормушками сахарозы.Пространственная предвзятость выбора пчелы (например, из-за расстояния до входа в улей или направления солнца) устранялась с помощью различных вариантов размещения рациона. Кроме того, эффекты пространственного смещения были смягчены поворотом кормовых площадок на 90 ° после записи посещения пчел каждый час (см. Раздел «Данные о посещениях пчел, собранных диетических весах и общей активности колоний»).
Дни сбора данных
Семейства медоносных пчел разместили в палатках 4 ноября, после захода солнца. Через 4 дня эксперимент был завершен с общим воздействием кормушек на каждую колонию в 19:15 ч, в среднем 4:45 ч в день (5, 6, 7 и 8 ноября).Семьи шмелей были протестированы через месяц и помещены в палатки 4 декабря на закате. Эксперимент со шмелями был завершен после того, как 20 из 24 семей шмелей продемонстрировали активность в кормлении на арене кормления (≥1 пчела). Семейства шмелей показали незначительную активность в течение первых 5 дней (исключая), включая 2 дня с дождем и низкими температурами. Воздействие кормушек на каждую колонию шмелей в последующие дни составляло 26:00 часов, в среднем 5:15 часов в день (10, 11, 12, 13 и 14 декабря).Средняя дневная температура во время эксперимента с медоносной пчелой была на 2,8 ° C выше, чем в эксперименте с шмелем (Приложение 1; t = 2,60, p = 0,04 * ), но высокие дневные температуры были благоприятны для колоний в качестве корма, и не было обнаружено различий между испытаниями на медоносных пчелах и шмелях (в среднем 24,2 ° C; t = 2,02, df = 7, p = 0,08).
Питатели для раствора сахара
Колонии были размещены в туннелях и снабжены питателями (внешними по отношению к колониям), содержащими сахарные растворы в пяти квадратных пластиковых контейнерах объемом 1 л, вмещающих 400 г раствора на дозатор.У каждого раствора сверху был квадратный поплавок с пузырчатой пленкой, служивший платформой для приземления и питья пчелами. Растворы хранили ad libitum и обновляли каждый день или второй день (закрывая кормушки крышками на ночь). Путем растворения сахара в воде были получены пять растворов с градиентом концентрации 16,5, 18,5, 20,5, 22,5, 24,5% (мас. / Мас.) Сахарозы, что подтверждено рефрактометрией. Этот диапазон соответствует концентрации нектара на нашей пасеке в течение ноября и составляет 20,7% ± 1,6 SEM ( n = 22 отобранных сборщика нектара) (Тамар Дрезнер-Леви, неопубликованные данные, 2003).
Питатели пыльцы
Колонии были снабжены кормушками (внешними по отношению к колониям), содержащими 100 г диетических партий пыльцы в пяти чашках Петри диаметром 13 см (20 г диеты на чашку). Эти количества обновлялись каждый день или второй день (закрытие посуды на ночь) при условии ad libitum , чтобы гарантировать отсутствие ограничений на варианты выбора. Каждый рацион содержал 70% измельченной смеси гранул пыльцы, собранной пчелами, с 30% смесью различных соотношений тапиоки к казеину (10: 0, 9: 1, 8: 2, 7: 3 или 6: 4).Казеин часто используется для изменения уровня белка в тестовых диетах (Pirk et al., 2010; Corby-Harris et al., 2018). Мука тапиоки использовалась в качестве наполнителя для создания разбавленных казеиновых белков (Hendriksma and Shafir, 2016).
Три диетических компонента (тапиока, казеин и пыльца) смешивали в настольном смесителе, и полученные диеты пропускали через сито для меда и хранили в холодильнике до использования. Тапиока покупалась как пищевая мука (Дюшан, Ган-Шмуэль, Израиль). Казеин (C7078, Sigma, Миссури, США) сначала измельчали в шаровой мельнице (Pulverisette 6, Fritsch GmbH, Идар-Оберштайн, Германия), применяя 7 шариков за 2 цикла по 3 мин при 300 об / мин для партий 80 г казеина. .Пыльца собиралась с помощью ловушек на местных ульях медоносных пчел в течение года и хранилась замороженной при -20 ° C. Эти смешанные гранулы цветочной пыльцы (5,9 кг) гомогенизировали с помощью промышленного смесителя.
Анализ белков во всех диетах и пищевых компонентах с пыльцой
Чтобы изучить потребление макронутриентов колонией в геометрической структуре питания, были определены абсолютные количества белка (P) и углеводов (C) в корме. Концентрации пищевого белка оценивали с помощью анализа азота по Кьельдалю (Foss Tecator Kjeltec Auto Sampler System 1035 Analyzer, Хиллерод, Дания).Образцы 200 мг окисляли H 2 SO 4 с катализаторами Se и CuSO 4 при 400 ° C. N-амины, которые были преобразованы в (NH 4 ) 2 SO 4 , в дальнейшем титровались HCl для определения количества азота. % Сырого протеина (мас. / Мас.) Рассчитывали как% N, умноженный на коэффициент пересчета 6,25 (Roulston and Cane, 2000) или 6,38 для казеина (лист технических данных Sigma).
Были приготовлены отдельные партии рациона для испытаний медоносных пчел и шмелей с использованием одной и той же смеси гранул пыльцы, но с использованием разных партий казеина и тапиоки, что, возможно, объясняет небольшие различия (1.В среднем 9%) по содержанию белка. Конечный рацион пыльцы содержал 17,3, 18,1, 19,9, 22,3 и 24,6% белка в опыте с медоносными пчелами и 14,5 16,4, 19,0, 22,2 и 21,8% белка в опыте с шмелями. Все образцы были измерены в двух экземплярах.
Данные о посещениях пчел, собранных диетических весах и общей активности колоний
Пчелы, собирающие пыльцу или сахарозу, подсчитывали с помощью цифровой фотографии приблизительно с часовыми интервалами. Количество собранного рациона измерялось с точностью до нуля.01 г пыльцы и 1 г раствора сахарозы. Все кормушки взвешивали до и после воздействия на колонию. Данные по колониям, не добывающим пищу (всего 0–2 зарегистрированных посещения), показали среднесуточное усыхание пыльцевого рациона на 1,5% и испарение растворов сахарозы на 2,8%, для чего были скорректированы данные о массе корма в активных колониях.
Отсутствие кормодобывания считалось признаком бездействия колонии. Колонии считались неактивными, если за весь период эксперимента на кормовой арене насчитывалось не более 7 собирателей.Колонии, которые были неактивны для сбора сахарозы ( N = 12), имели среднее 0,02 ± 0,01 SE собирателей сахарозы, подсчитываемых за одно наблюдение, и собирали среднее значение 0,00 г / ч -1 ± 0,16 SE раствора сахарозы, т. Е. Незначительно отличалось от нуля. (по одной выборке t -тест; t = 0,0, p = 1,0). Колонии, неактивные в сборе пыльцы ( N = 28), имели в среднем 0,09 ± 0,02 SE собирателей пыльцы, а собранные средние 0,01 gh −1 ± 0,02 SE пыльцевой диеты, также незначительно отличавшейся от нуля (одна выборка t -тест; t = 0.69, p = 0,49).
Для всех активных колоний относительный набор веса рациона рассчитывали путем деления суммы собранных весов на уровень концентрации на общий собранный вес. Кроме того, был рассчитан индекс кормовой нагрузки для каждого уровня концентрации путем деления веса рациона, собранного за час, на среднее количество пчел на рационе (единица: –1 · пчела –1 ). Предполагая, что продолжительность времени, которое пчела проводит на кормушке, пропорциональна количеству собранного ею корма и вероятности того, что она будет запечатлена при переписи фотографий, индексы нагрузки являются приближенными к нагрузкам культур нектара, зависящим от концентрации в рационе, и нагрузкам корбикулярной пыльцы. , соответственно.Идентификационные данные отдельных пчел не регистрировались, однако наши данные все же могут дать представление об индивидуальных решениях о кормлении. Собиратели могут по-разному проводить время на поисках корма между разными кормушками, поэтому их выбор, относительное количество удаляемой пищи, а также кормовая нагрузка могут служить показателями индивидуальной мотивации пчел относительно различных концентраций питательных веществ в предлагаемых рационах.
P: C Коэффициент усвоения рациона активными колониями
СоотношениеP: C в колониях рассчитывали путем деления граммов поглощения P на поглощение C.Неактивные колонии были исключены (собрано ≤7 собирателей или ≤0 г корма). Потребление P было суммой веса собранной с пищей пыльцы, умноженной на% P. Потребление C колониями представляло собой сумму собранных диетических весов пыльцы, умноженных на% C в пыльцевом рационе, плюс вес собранного раствора сахарозы, умноженный на% C в растворе. Мы предположили, что гранулы пыльцы содержат 50% углерода (Nicolson, 2011), который включает крахмал пыльцы (0–22%; Roulston and Buchmann, 2000), пыльцевые сахара (9,6%; Speranza et al., 1997) и нектарные сахара, добавляемые собирателями при осаждении пыльцы (Bertoncelj et al., 2018). Считалось, что казеин (спецификация продукта, Sigma) и тапиоковая мука (пищевой лейбл, Duchan) содержат 0 и 99,8% углерода соответственно. Таким образом, было подсчитано, что пять пыльцевых рационов содержат 64,2, 61,3, 58,4, 55,5 и 52,6% углерода соответственно.
Статистика
Анализы проводились с помощью статистического программного обеспечения JMP 13.1 Pro.
Активность колонии
Число активных и неактивных семей медоносных пчел и шмелей сравнивалось с помощью точных тестов Fisher в отношении сбора пыльцы, сбора сахарозы и общей активности кормодобывания (Таблица 1).
Подсчет пчел
Обобщенные линейные модели (GLM) были подогнаны по максимальной вероятности с экспоненциальным распределением и функцией взаимной связи для анализа воздействия обработки на количество собирателей. Количество собирателей пыльцы было проанализировано на эффекты лечения белком (3 уровня P– / P / P + для медоносных пчел; 2 уровня P / P + для шмелей), а количество собирателей нектара на эффекты лечения углеводами (2 уровня C– / C для медоносных пчел) ; 2 уровня C / C + для шмелей). Подгонка модели оценивалась с помощью теста Пирсона χ 2 на избыточную дисперсию.Окончательные приближения значений p были основаны на тестах Вальда.
Распределение собирателей по рационам, лечебным средствам и типам пчел
Процент сборщиков пыльцы от общего числа сборщиков был принят как мера относительного распределения сборщиков пыльцы. Распределение (% данных, преобразованных в квадратный корень арксинуса) в качестве переменной ответа было проанализировано с помощью однофакторного дисперсионного анализа с обработкой в качестве независимой переменной (3 уровня) отдельно для медоносных пчел и шмелей. Апостериорные сравнения проводили с помощью тестов Тьюки на честную значительную разницу (HSD).Относительное выделение кормильцами семей медоносных пчел и шмелей сравнивали при обработке PC (то есть в базовых семьях) с типом пчелы в качестве независимой переменной (2 уровня).
Соотношение P: C
Потребление белка и углеводов колониями описывалось для каждой обработки как отношение P к C. Для исходных колоний (обработка PC) соотношения P: C на колонию сравнивали среди типов пчел (2 уровня) с помощью однофакторного дисперсионного анализа по рангам (критерий Краскала-Уоллиса). Неосновные обработки колоний (2 уровня) сравнивали отдельно для медоносных пчел и шмелей.
Влияние реакции на дозу при сборе диеты и переносе нагрузки
Относительные собранные веса рациона и индексы кормовой нагрузки сравнивались с помощью ANCOVA отдельно для медоносной пчелы и шмеля с независимыми переменными концентрацией в рационе (непрерывная переменная) и обработкой (3 уровня). Значительные взаимодействия могут указывать на то, что выбор пчел в зависимости от концентрации различается из-за манипуляций с колонией. Для исходных семей (обработка PC) относительные веса и индексы нагрузки сравнивались с независимыми переменными концентрацией в рационе (непрерывная переменная) и типом пчелы (2 уровня).Здесь взаимодействие покажет разные реакции выбора медоносных пчел и шмелей в отношении концентраций фосфора или углерода в рационе. Незначительные взаимодействия ( p ≥ 0,05) были удалены из моделей. Поскольку пропорциональные данные в среднем идентичны между группами ( p = 1,0), ковариант (т.е. лечение или тип пчелы) также был удален из модели в тех случаях, когда взаимодействие не было значимым.
Результаты
Уровень колонии медоносных пчел, уравновешивающий усвоение фосфора и углерода
Всего было зарегистрировано 5 149 посещений медоносными пчелами на диетах.В целом семьи медоносных пчел ( N = 24) собрали 493 г пищевой пыльцы и 15,7 кг раствора сахарозы (соотношение 1:32). Колонии были значительно более активными в отношении сбора сахарозы по сравнению со сбором пыльцы (Fisher Exact p <0,001). Подсчеты собирателей пыльцы показали разницу при манипуляциях с белками (рис. 1A; N = 12, χ 2 = 13,1, df = 2, p = 0,001). Сборщики пыльцы были значительно более активны при обработке P по сравнению с обработкой P (χ 2 = 5.27, df = 1, p = 0,02) и P + (χ 2 = 10,6, df = 1, p = 0,001). Количество собирателей пыльцы не различалось между колониями, обработанными P и P + (χ 2 = 2,53, df = 1, p = 0,11). Количество собирателей нектара не различалось между колониями в зависимости от обработки C (сравнение C и C–, Рисунок 1A; χ 2 = 2,55, df = 1, p = 0,11).
Рисунок 1 .Эффекты на уровне колоний от манипуляций с питательными веществами в улье. Количество собирателей нектара (NF) и количество собирателей пыльцы (PF) показано в (A) для медоносных пчел (HB) и в (D) для шмелей (BB) в условиях повышенного или пониженного содержания белка (P ) или углеводные (C) продовольственные магазины. Показаны средние значения активных колоний подсчетов собирателей ± шкала ошибок SE, с пунктирными диагоналями, указывающими среднее число собирателей, подсчитанное в базовых колониях. Статистическая значимость обозначена звездочкой (*).Распределение кормов для нектара (N) и пыльцы (P) показано в (B, E) (средние значения в% ± шкала ошибок SE), при этом значимые эффекты обработки обозначены разными буквами ( post hoc Tukey HSD). Отношения уровня колоний P: C указаны в (C, F) пунктирными линиями, показывающими среднее потребление белка к углеводам ± шкала погрешности SE для обработки. Post hoc Статистическая значимость отмечена звездочкой (*).
В условиях избытка пыльцы и уменьшения количества углеводов (P + C–) медоносные пчелы перешли к разделению труда при добывании пищи, основанному на С, с большим количеством собирателей сахарозы (Таблица 2).Кроме того, относительное сравнение распределения собирателей показало эффекты обработки [Рисунок 1B; N = 24, F (2, 21) = 4,33, p = 0,027], с наименьшим количеством кормов пыльцы при обработке P +. Эти колонии показали относительно высокий уровень потребления сахарозы (рис. 1B; 97% нектара по сравнению с 3% сборами пыльцы, что проиллюстрировано их замечательным соотношением потребления P: C 1/2080 (рис. 1C). Для обратной обработки депривации пыльцы и углеводов). (P – C–), медоносные пчелы перенесли свои задачи на сбор пыльцы (Таблица 2).Почти половина колонии собирала пыльцу (Рисунок 1B; 46% пыльцы против 56% сахарозы), при этом общее потребление P-C сдвинуто до 1:14 (Рисунок 1C). Соотношения P: C в семьях медоносных пчел, не получавших исходной обработки, значительно различались [P + C– против P – C +; F (1, 14) = 8,09, p = 0,013].
Таблица 2 . Количество активных и неактивных колоний.
Уровень колонии шмелей уравновешивает поглощение фосфора и углерода
Всего было зарегистрировано 714 визитов шмелей на диеты.В целом семьи шмелей ( N = 24) собрали в общей сложности 155 г пыльцы корма и 2,0 кг раствора сахарозы (соотношение 1:13). Для семей шмелей активности по сбору сахарозы и пыльцы существенно не различались (Таблица 2; N = 12 против . N = 8, Fisher Exact p = 0,38) (Таблица 2). Подсчет собирателей пыльцы показал отсутствие разницы между манипуляциями P и P + (Рисунок 1D; N = 8, χ 2 = 3,56, df = 1, p = 0.06), однако количество собирателей нектара было значительно ниже при обработке C + по сравнению с обработкой C (Рисунок 1D; N = 12, χ 2 = 8,32, df = 1, p = 0,004).
Колонии шмелей ( N = 13) продемонстрировали значительное влияние обработки на распределение кормодобывающих средств [Рисунок 1E; F (2, 10) = 14,2, p = 0,001]. Примечательно, что колонии, обработанные PC +, показали значительно более низкое распределение рабочих для сбора сахарозы по сравнению с двумя другими обработками (рис. 1E; 9% сбора сахарозы vs.91% для пыльцы). Колонии P + C имели самое низкое соотношение P: C, то есть 1:51, что контрастировало с обратной обработкой P + C, где колонии потребляли соотношение PC + 1: 5 (рис. 1F). Отношения P: C в колониях, отличных от исходного уровня, значительно отличались друг от друга [P + C vs. PC +; F (1, 5) = 675,8, p <0,001].
Различия в уровне колоний между видами пчел
Значительные различия между видами пчел и внутри них наблюдались в отношении активности колоний (Таблица 2).Колонии медоносных пчел были более активными, чем колонии шмелей (общее количество собирающих корм 24> 13 колоний, соответственно, Fisher Exact p <0,001; кормление сахарозой 24> 12 колоний, соответственно, Fisher Exact p <0,001; сбор пыльцы 12 ≈ 8 колоний соответственно, Fisher Exact p = 0,38). На уровне колонии медоносные пчелы, но не шмели, уравновешивали добычу пыльцы в ответ на потребности колонии, и, напротив, шмели, но не медоносные пчелы, уравновешивали свой сбор сахарозы (Рисунки 1A, D).Обработка ПК (исходные колонии) показала, что выделение 21,1% сборщиков пыльцы для семей медоносных пчел ( N = 8) не отличалось от выделения 21,9% сборщиков пыльцы для семей шмелей ( N = 6) [центральные столбцы Фигуры 1Б, Д; F (1, 12) = 0,05, p = 0,83]. Базовые колонии обоих видов пчел не различались по соотношению потребления белка и углеводов, с соотношением P: C 1:28 против 1:17, соответственно [Рисунки 1C, F; F (1, 11) = 0.09, p. = 0,77].
Индивидуальные реакции медоносных пчел на потребности в питательных веществах в фосфорилированных и углеродных клетках
В целом, медоносные пчелы собрали в 23 раза больше сахарозы самой высокой концентрации по сравнению с самой низкой концентрацией (42,0 против 1,8%, соответственно), демонстрируя весьма значимое предпочтение более высоких концентраций C [Рисунки 2A – C, концентрация; F (1, 109) = 205,5, p <0,001]. Пчелы в ульях с добавлением пыльцы (P + C–) показали менее выраженное предпочтение по сравнению с двумя другими видами обработки, а именно в 7 раз больше собирали раствор сахарозы с самой высокой концентрацией по сравнению с самой низкой концентрацией, причем это различие иллюстрируется более пологим наклоном [Рисунок 2A vs.Рисунок 2Б, В; наклонное взаимодействие F (2, 109) = 3,69, p = 0,028]. Кормовые нагрузки сахарозы также были значительно выше, когда пчелы посещали кормушки с более высоким содержанием углерода [концентрация; F (1, 111) = 46,1, p <0,001] (Рисунки 2G – I). Сборщики не показали зависимого от концентрации эффекта на нагрузку в ответ на манипуляции с колониями P и C, то есть сходные наклоны в трех группах обработки [Рисунки 2G – I; Взаимодействие наклона F (2, 109) = 2.94, p. = 0,057]. На загрузку раствора сахарозы не влияли манипуляции с колониями [обработка; F (2, 111) = 1,61, p = 0,20].
Рисунок 2 . Предпочтение медоносных пчел и шмелей при сборе сахарозы. С концентрацией углеводов сахарозы в рационе в процентах на оси x, относительный сбор сахарозы отдельными пчелами показан в виде весовых пропорций на оси y (A – F) и нагрузки сахарозы на оси y (G– Л) .Линии тренда показывают значимость доза-ответ для каждого лечения, включая манипуляции с белками (P) и углеводами (C), в колониях медоносных пчел (слева) и колониях шмелей (справа).
Предпочтение концентрацииP пчелами-собирателями различается в зависимости от обработки колонии [влияние наклона; F (2, 49) = 6,31, p = 0,004]. В колониях, обработанных PC и P – C–, фуражиры предпочитали собирать рационы пыльцы с более низким содержанием P (казеина) [Рисунки 3B, C; Ф. (1, 18) = 64.2, p <0,001 и F (1, 23) = 77,47, p <0,001, соответственно]. Это отрицательное предпочтение P было значительно сильнее при лечении PC по сравнению с обработкой P – C– [наклон взаимодействия; F (1, 41) = 18,1, p <0,001]. Не было никаких указаний на отрицательное предпочтение P в колониях P + C– [Рисунок 3A; F (1, 8) = 0,17 , p = 0,69]. Напротив, при различных концентрациях обработка не влияла на содержание пыльцы [Рисунки 3G – I; Взаимодействие наклона F (2, 47) = 0.50, p. = 0,61]. В целом, на содержание пыльцы медоносных пчел не влияла концентрация белка [ F (1, 49) = 0,14, p = 0,71]. Пыльцевая нагрузка действительно различалась в зависимости от обработки P + C–, PC и P – C– [ F (2, 49) = 7,56, p = 0,001], при этом исходные колонии-фуражиры (рис. 3H) демонстрируют более высокая нагрузка по сравнению с пополненными пыльцой колониями-сборщиками (P + C– Рисунок 3G).
Рисунок 3 . Предпочтение медоносных пчел и шмелей во время сбора пыльцы.С концентрацией белка пыльцы в рационе в процентах на оси x, относительный сбор пыльцы отдельными пчелами показан в виде весовых пропорций на оси y (A – F) и нагрузки пыльцы на оси y (G– Л) . Линии тренда показывают значимость доза-ответ для каждого лечения, включая манипуляции с белками (P) и углеводами (C), в колониях медоносных пчел (слева) и колониях шмелей (справа). Полосатые линии тренда указывают на значительную доза-ответную реакцию для шмелей в целом, хотя этот эффект был незначительным для групп лечения индивидуально.
Индивидуальные реакции шмелей на потребности в питательных веществах P и C
Шмели-фуражиры при обработке PC + не поддерживали высокие концентрации C [Рисунок 2F; F (1, 3) = 3,55, p = 0,16]. Это было значительным контрастом [наклон взаимодействия; F (2, 49) = 19,5, p <0,001] с собирателями колоний, обработанных P + C и PC, которые действительно показали значительное предпочтение более высоким концентрациям сахарозы [Рисунок 2D, E; Ф. (1, 23) = 68.6, p <0,001 и F (1, 23) = 48,3, p <0,001, соответственно] и собрали пропорционально в 8 раз больше сахарозы самой высокой концентрации по сравнению с самой низкой концентрацией. Пищевые нагрузки шмелей различались по зависимости доза-ответ C [наклон взаимодействия F (2, 45) = 15,6, p <0,001]. При обработке PC собиратели увеличили загрузку урожая с более высокой концентрацией C [Рисунок 2K; Ф. (1, 21) = 7.56, p = 0,012], но такой паттерн не наблюдался в колониях, обработанных P + C и PC + [Фигуры 2J, L; F (1, 22) = 3,18, p = 0,09 и F (1, 2) = 2,87, p = 0,23, соответственно]. Люди собирали более легкие нагрузки при лечении PC по сравнению с обработками P + C и PC + [Рисунки 2J – L; F (2, 45) = 7,40, p = 0,002, post-hoc A B A]. Эти данные предполагают, что добавление пирожков с пыльцой побудило пчел собирать больше сахарозы (рис. 2J), что может быть связано с повышенной производительностью расплода, мотивируя более высокое потребление топлива (Приложение 1).Примечательно, что, давая шмелям в улье раствор сахарозы, собиратели загружали в основном наименее концентрированный раствор (например, 18,4 г / ч −1 · посещение −1 ), что позволяет предположить, что собиратели перешли к поведению сбора воды (рис. 2L).
Шмели, по сравнению со всеми рассматриваемыми обработками, показали значительное предпочтение в отношении сбора рационов с более низкими концентрациями белка [Рисунки 3D – F; F (1, 31) = 7,93, p = 0,008]. На это предпочтение, однако, не повлияли манипуляции с колониями P или C [наклонное взаимодействие; F (2, 34) = 0.62, p = 0,54 и лечение F (2, 34) = 0,00, p = 1,00]. Также не было никакого влияния обработки на количество пыльцы-фуражиры [Рисунки 3J – L, взаимодействие на склоне; F (2, 31) = 0,59, p = 0,56]. На содержание пыльцы шмелей не влияла концентрация белка [ F (1, 33) = 0,60, p = 0,44], а количество кормов не различается в ответ на обработку колоний P и C [Рисунки 3J – L ; F (2, 33) = 0.25, p. = 0,78].
Различия индивидуальной реакции между видами пчел
Прямое сравнение было проведено для семей медоносных пчел и шмелей только для одной обработки, при которой они находились в том же состоянии, что и при исходной обработке PC (Таблица 2). Оба вида предпочитали собирать больше раствора сахарозы с более высокими концентрациями C [Рисунки 2B, E; F (1, 58) = 104,6, p <0,001], и этот эффект был одинаковым для разных видов [наклонное взаимодействие; Ф. (1, 56) = 1.07, p = 0,31, а тип пчелы F (2, 34) = 0,00, p = 1,00]. Медоносные пчелы и шмели увеличивали нагрузку за счет более высоких концентраций сахарозы [ F (1, 55) = 21,2, p <0,001], без разницы между видами в их ответной концентрации [наклонное взаимодействие; F (1, 54) = 0,00, p = 0,96]. Сборщики шмелей показали в 3 раза более высокое содержание сахарозы по сравнению с сборщиками медоносных пчел [пчелиный тип; Ф. (1, 55) = 27.2, p. <0,001].
Медоносные пчелы и шмели-фуражиры предпочитали корм для сбора пыльцы с более низкой концентрацией белка [Рисунки 3B, E; F (1, 31) = 41,9, p <0,001], но это предпочтение было сильнее у медоносных пчел по сравнению с шмелями [Рисунки 3B, E, взаимодействие на склоне; F (1, 31) = 24,0, p <0,001]. Также было в 3 раза выше содержание пыльцы у собирателей шмелей по сравнению с собирателями медоносных пчел [тип пчелы; Ф. (1, 32) = 8.24, p. = 0,007]. Различия в реакции на дозу белка при загрузке не было обнаружено между видами [наклонное взаимодействие; F (1, 31) = 0,96, p = 0,33], и концентрация белка не была обнаружена на нагрузку [ F (1, 32) = 0,28, p = 0,60].
Обсуждение
В этом исследовании были изучены два очень разных вида эусоциальных пчел ( медоносных пчел A. mellifera, и шмелей B. terrestris, ) в ответ на изменение запасов углеводов и белков в их колониях.При исходной обработке P-C соотношение P-C-фуражиров и собранных P: C-кормов было поразительно схожим между двумя видами, с почти 80% -ным распределением C при кормлении (Рисунки 1B, E) и примерно сопоставимыми соотношениями P: C. собраны (Рисунки 1C, F). В целом, результаты подтверждают исходную гипотезу о том, что оба вида пчел сместят разделение труда, чтобы противодействовать сдвигам в балансе питательных веществ в колониях.
Например, семьи медоносных пчел и шмелей с нарушенным балансом P: C произвели адаптивные изменения в соотношении P: C собранного корма (Рисунки 1C, F).Таким образом, разделение труда для обоих видов повлияло на фактическую массу питательных веществ, потребляемых собирателями. В целом, эти данные показывают, что оба вида обладают способностью изменять усилия по добыче пищи на уровне колоний, чтобы сбалансировать потребление белков и углеводов.
Существовали также некоторые ключевые различия между видами в их специфической реакции на энергетический баланс колоний (суммированные в Таблице 3). Колонии медоносных пчел и шмелей изменили количество собирателей и процент посещений кормов (рис. 1A, B, D, E) в ответ на манипуляции с питательными веществами.Однако сдвиги были значительными только для обработок со сдвигом P у медоносных пчел и значительными только для обработок со сдвигом C у шмелей. Эти данные показывают, что медоносные пчелы более чувствительны к потребностям фосфора, тогда как шмели более чувствительны к потребностям в фосфоре. Это согласуется с известной высокой чувствительностью семей медоносных пчел к потребностям колонии в пыльце (Camazine, 1993), а также согласуется с ранее опубликованными углеводными ответами у трех других видов шмелей (Cartar, 1992). Различная динамика между видами пчел согласуется с их разным жизненным циклом.Ожидается, что колонии многолетних медоносных пчел, которые точно настроены на накопление большого количества меда, необходимого для перезимовки, всегда будут выделять собирателей для сбора нектара, при этом более точно регулируя сбор пыльцы в соответствии с запасами колонии и текущими потребностями в расплодах (Keller et al. , 2005). Ежегодные колонии шмелей, которые не хранят большое количество меда для зимовки, должны сокращать выделение собирателей нектара, как только будут достигнуты достаточные запасы углеводов, но всегда выделяют фуражиров для сбора пыльцы для поддержания максимальной продуктивности расплода (Cartar, 1992).
Таблица 3 . Краткое изложение эффектов лечения, концентрации в рационе, видов пчел и взаимодействий.
В целом предсказывалось, что предпочтения отдельных пчел будут смещены в сторону более высоких концентраций питательных веществ во время кормодобывания. В соответствии с этим предсказанием, собиратели обоих видов обычно предпочитали кормушки с более высокой концентрацией сахарозы (рис. 2, верхний ряд). Основной ключевой вопрос заключался в том, изменились ли предпочтения собирателей в концентрации в условиях депривации колонии по сравнению с насыщением.Было обнаружено, что оба вида пчел изменяют индивидуальную реакцию при изменении уровня питательных веществ в колонии. Что касается медоносных пчел, предпочтение сахарозы снижалось в колониях с добавлением пыльцы. Этот сдвиг в сторону принятия более низких концентраций мог быть вызван сенсорной модуляцией в зависимости от состояния питания отдельной пчелы (рис. 2А). Однако причиной также может быть сдвиг в распределении пыльцы по сравнению с собирателями нектара. Вероятно, что среди наблюдаемых 97% сборщиков нектара были пчелы с предрасположенностью к сбору пыльцы (рис. 1B).У собирателей пыльцы более низкий порог чувствительности к сахарозе (Pankiw, Page, 2000; Scheiner et al., 2001; Pankiw, 2003; Drezner-Levy et al., 2009). Следовательно, состояние питания колоний могло изменить распределение собирателей, а вместе с тем и среднюю сенсорную модальность предпочтения сахарозы.
Шмели-фуражиры также предпочитают более высокие концентрации сахарозы, за исключением обработки с добавлением сахарозы (рис. 2F). Здесь фуражиры показали заметный сдвиг в предпочтении низких концентраций сахарозы.С кормушкой в улье, содержащей 50% мас. / Об. Сахарозы, фуражиры, возможно, перешли к сбору кормов с высоким содержанием воды, даже при наличии дополнительных кормушек на экспериментальной арене (рис. S2).
Собиратели обоих видов обычно загружают больше сахарозы из кормушек с более высокой концентрацией (рис. 2, нижний ряд). Увеличение нагрузки с увеличением концентрации сахарозы согласуется с выводами Нуньеса (1966) и описательной моделью Варжу и Нуньеса (1991), в которых концентрация сахарозы является основным определяющим фактором нагрузки сельскохозяйственных культур.Однако для медоносных пчел характер нагрузки в ответ на концентрацию сахарозы не различается при лечении баланса питательных веществ в колониях (Рисунки 2G – I). Это говорит о том, что, несмотря на изменение потребностей колонии, отдельные пчелы поддерживают поведение, которое оптимизирует потребление энергии углерода (Schmid-Hempel et al., 1985; Kacelnik et al., 1986; Schmid-Hempel, 1987; Varjú and Núñez, 1991; Afik and Shafir. , 2007). В отличие от медоносных пчел, шмели продемонстрировали значительное влияние обработки колоний питательными веществами на среднюю нагрузку сахарозы (Рисунки 2J – L).В отличие от собирателей медоносных пчел, которые все одинаковы по размеру, шмели сильно различаются по размеру в пределах колонии (рис. S2), при этом фуражиры в среднем крупнее гнездовых пчел (Goulson et al., 2002). Одно из объяснений того, почему средняя нагрузка урожая шмелей была значительно ниже при обработке PC (рис. 2K), по сравнению с другими обработками, заключается в том, что общее количество собирателей было наибольшим при обработке PC. Из-за высокой потребности в фуражах как на пыльцу, так и на нектар (рис. 1D), это могло вызвать дополнительный набор фуражиров, в том числе шмелей меньшего размера.Другое объяснение состоит в том, что истощение питательных веществ привело к гибели пчел-собирателей (Дополнение 1), что могло (опять же) вызвать пополнение более мелких особей с более низкими урожаями (Hagbery and Nieh, 2012). Фактически, в то время как сборщики шмелей предпочитают более крупных особей, когда сборщики кормов истощаются из-за индуцированной смертности, вероятность перехода пчел в гнездо на добычу корма не зависит от их размера (Crall et al., 2018).
Индивидуальный уровень реакции на белковый корм резко отличался от такового на углеводный корм.Несмотря на то, что предпочтения отдельных фуражиров, согласно прогнозам, будут смещены в сторону более высоких концентраций питательных веществ P, вместо этого фуражи проявили отвращение к высоким концентрациям белка (рис. 3; верхний ряд). Почему пчелы избегали более высокой концентрации белка? Содержание пыльцы (70%) было постоянным для всех диет, поэтому эффект можно отнести к казеину или тапиоке в рационе. Либо пчелы обнаружили отвращение к казеину, либо пчелы были привлечены тапиокой. Когда тапиока всасывается, она может вызывать предпочтение, потому что крахмал гидролизуется в сахар α-амилазой, ферментом, вырабатываемым слюнными железами медоносных пчел (Wright et al., 2018). Впоследствии сладость может служить наградой и вызывать предпочтение. Жирность может быть еще одним объяснением того, почему пчелы могут испытывать отвращение к казеину. Технический казеин из коровьего молока имеет некоторое содержание жира (C7078, Sigma), и Хендриксма и Шафир (2016; добавка внутри) обнаружили, что более высокое содержание жиров в рационе отрицательно влияет на предпочтения пчел в рационе.
Несмотря на отталкивающий характер высоких концентраций протеина, сборщики медоносных пчел показали модуляцию индивидуальной реакции на протеин в зависимости от баланса питательных веществ в колонии.В исходных колониях собиратели показали наиболее выраженное отрицательное предпочтение концентрации, в то время как собиратели из пополненных пыльцой колоний были относительно неизбирательными. Это говорит о том, что сытые медоносные пчелы потеряли способность различать питательные вещества и / или предпочтение (рис. 3А).
Пыльцевая нагрузка (рис. 3, нижний ряд) была выше у более крупных шмелей, чем у медоносных пчел, что также было обнаружено в естественных условиях (Minahan and Brunet, 2018). У медоносных пчел наблюдался значительный лечебный эффект: колонии, содержащие пыльцу, показали меньшую нагрузку по сравнению с исходным уровнем.Таким образом, медоносные пчелы, по-видимому, модулируют свои реакции кормления P в ответ на сдвиги энергетического баланса колонии. Шмели продемонстрировали менее выраженное отвращение к содержанию белков пыльцы в пище, и не было никаких значительных эффектов лечения (Рисунки 3D – F), то же самое верно и в отношении содержания пыльцы (Рисунки 3J, K, L). Это говорит о том, что шмели менее восприимчивы к концентрации белка в рационе и / или что они кормят корм в соответствии с заданными базовыми предпочтениями, на которые не влияют различия в потребностях колоний.
Важно отметить, что наблюдаемые сдвиги в ответах на индивидуальном уровне (таблица 3) могут не отражать изменения поведения индивидов; скорее они могут представлять изменения в распределении фуражиров, которые различаются по размеру и / или пороговой чувствительности к пищевым стимулам. Будущие эксперименты с отслеживанием отдельных пчел до и после смены питательных веществ в колониях необходимы, чтобы разобраться, представляют ли наблюдаемые различия прямые индивидуальные сдвиги или сдвиги в распределении колоний у особей с различными предпочтениями в питательных веществах.Кроме того, будущие исследования индивидуальной реакции и реакции колонии для колоний на разных фазах жизненного цикла колонии дополнят текущие результаты, поскольку ответы собирателей могут измениться, когда колонии готовятся к воспроизводству или достигают конца сезона (Pankiw and Page, 1999; Johnson , 2010).
Возвращаясь к исходным гипотезам, в этом исследовании представлены доказательства, подтверждающие гипотезу о том, что как высокоэусоциальные, так и примитивно эусоциальные пчелы уравновешивают потребности колонии в питательных веществах за счет смещения разделения труда на добычу пищи.Хотя шмели не так высоки социальны, как медоносные пчелы, тот факт, что они преодолели «порог суперорганизма» по некоторым определениям (потому что у них постоянные морфологические касты; Boomsma and Gawne, 2018), указывает на то, что принятие решений на уровне колонии — хорошее развились даже в «более простых» сообществах социальных насекомых. Основываясь на различиях в уровне социальности между медоносными пчелами и шмелями, мы также предсказали более сильные индивидуальные реакции у шмелей по сравнению с медоносными пчелами. Вопреки этому прогнозу, мы нашли доказательства сдвигов в индивидуальной восприимчивости к питательным веществам (особенно в отношении предпочтения углеводов и их загрузки) для обоих видов.Вместо того, чтобы быть связанными с различиями в социальности, различия между видами в их специфических реакциях (суммированных в Таблице 3) вместо этого могут быть связаны с различиями в особенностях жизненного цикла колоний (таких как уровень накопления пищи, форма обеспечения расплода и т. Д.). Таким образом, мы призываем к более тщательному учету особенностей жизненного цикла, таких как родительская забота, накопление запасов и сезонность, при принятии решений о поиске пищи и в контексте OFT.
Авторские взносы
Дизайн исследования, сбор данных и анализ данных были выполнены HH.Рукопись написана HH, AT и SS.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Мы благодарны Совету по исследованиям в области биотехнологии и биологических наук (BBSRC), Совету по исследованиям окружающей среды (NERC), Wellcome Trust, Департаменту окружающей среды, продовольствия и сельских районов (Defra) и правительству Шотландии в рамках инициативы по насекомым-опылителям грант №: BB / I000968 / 1) и Министерство сельского хозяйства США — Национальный институт продовольствия и сельского хозяйства (грант №: 2016-07965) для финансирования, а также проф.Доктор Амос Нусинович и Захи Рабиновиц за подготовку казеина, Самир Мабджиш за анализ белков и Хаим Калев за размещение колонии в его гранатовом саду.
Дополнительные материалы
Дополнительные материалы к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/articles/10.3389/fevo.2019.00177/full#supplementary-material
Список литературы
Афик, О., Шафир, С. (2007). Влияние температуры окружающей среды на загрузку урожая у медоносной пчелы, Apis mellifera (Hymenoptera: Apidae ). Энтомол. Gen. 29, 135–148. DOI: 10.1127 / entom.gen / 29/2007/135
CrossRef Полный текст | Google Scholar
Айзен, М. А., Смит-Рамирес, К., Моралес, К. Л., Виели, Л., Саез, А., Бараона-Сеговия, Р. М. и др. (2018). Скоординированная политика глобального ввоза видов необходима для сокращения серьезных вторжений во всем мире: случай чужеродных шмелей в Южной Америке. J. Appl. Ecol. 56, 100–106. DOI: 10.1111 / 1365-2664.13121
CrossRef Полный текст | Google Scholar
Алтай, с.З., Пирк, К. В., Крю, Р. М., и Николсон, С. В. (2010). Сближение целевых показателей потребления углеводов у пчел, содержащихся в клетках, которым кормили разные источники белка. J. Exp. Биол. 213, 3311–3318. DOI: 10.1242 / jeb.046953
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Амент, С. А., Ван, Ю., Робинсон, Г. Э. (2010). Пищевая регуляция разделения труда у медоносных пчел: к перспективе системной биологии. Wiley Interdiscip. Rev. Syst.Биол. Med. 2, 566–576. DOI: 10.1002 / WSBM.73
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бертонсель, Дж., Полак, Т., Пучихар, Т., Лилек, Н., Кандольф Боровшак, А., и Корошец, М. (2018). Углеводный состав пыльцы словенской пчелы. Внутр. J. Food Sci. Technol. 53, 1880–1888. DOI: 10.1111 / ijfs.13773
CrossRef Полный текст | Google Scholar
Бумсма, Дж. Дж., И Гоун, Р. (2018). Сверхорганизм и кастовая дифференциация как точки невозврата: как основные эволюционные переходы были потеряны при трансляции. Biol. Ред. 93, 28–54. DOI: 10.1111 / brv.12330
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бергетт Д. М. и Янг Р. Г. (1974). Запасы липидов медовыми муравьями пополняются. Ann. Энтомол. Soc. Являюсь. 67, 743–744. DOI: 10.1093 / aesa / 67.5.743
CrossRef Полный текст | Google Scholar
Камазин, С. (1993). Регулирование сбора пыльцы медоносными пчелами — как собиратели оценивают потребность колонии в пыльце. Behav. Ecol.Sociobiol. 32, 265–272. DOI: 10.1007 / BF00166516
CrossRef Полный текст | Google Scholar
Картар, Р. В. (1992). Регулировка усилий по добыче корма и переключение задач в управляемых энергией колониях диких шмелей. Anim. Behav. 44, 75–87. DOI: 10.1016 / S0003-3472 (05) 80757-2
CrossRef Полный текст | Google Scholar
Корби-Харрис, В., Снайдер, Л., Мидор, К., и Айотт, Т. (2018). Медсестры медоносной пчелы ( Apis mellifera ) не употребляют пыльцу из-за ее питательных качеств. PLoS ONE 13: e01
. DOI: 10.1371 / journal.pone.01
CrossRef Полный текст | Google Scholar
Кролл, Дж. Д., Гравиш, Н., Маунткасл, А. М., Кочер, С. Д., Оппенгеймер, Р. Л., Пирс, Н. Е. и др. (2018). Пространственная верность рабочих предсказывает коллективную реакцию на нарушение общественного порядка. Nat. Коммуна . 9: 1201. DOI: 10.1038 / s41467-018-03561-w
CrossRef Полный текст | Google Scholar
Дорнхаус А. и Читтка Л.(2001). Пищевая опасность у шмелей ( Bombus terrestris ): возможные механизмы и эволюционные последствия. Behav. Ecol. Sociobiol. 50, 570–576. DOI: 10.1007 / s002650100395
CrossRef Полный текст | Google Scholar
Дрезнер-Леви, Т., Смит, Б., и Шафир, С. (2009). Влияние кормовой специализации на различные учебные задачи медоносной пчелы ( Apis mellifera ). Behav. Ecol. Sociobiol. 64, 135–148. DOI: 10.1007 / s00265-009-0829-z
CrossRef Полный текст | Google Scholar
Галеф, Б.Дж. Младший, и Жиральдо, Л.А. (2001). Социальные влияния на добычу пищи у позвоночных: причинные механизмы и адаптивные функции. Anim. Behav. 61, 3–15. DOI: 10.1006 / anbe.2000.1557
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гоулсон Д. (2010). Шмели: поведение, экология и сохранение . Нью-Йорк, Нью-Йорк: Издательство Оксфордского университета.
Google Scholar
Гоулсон, Д., Пит, Дж., Стаут, Дж. К., Такер, Дж., Дарвилл, Б., Derwent, L.C., et al. (2002). Можно ли объяснить аллоэтизм у рабочих шмеля Bombus terrestris с точки зрения эффективности кормодобывания? Anim. Behav. 64, 123–130. DOI: 10.1006 / anbe.2002.3041
CrossRef Полный текст | Google Scholar
Хендриксма, Х. П., и Шафир, С. (2016). Сборщики меда уравновешивают дефицит питательных веществ в колониях. Behav. Ecol. Sociobiol. 70, 509–517. DOI: 10.1007 / s00265-016-2067-5
CrossRef Полный текст | Google Scholar
Качельник, А., Хьюстон, А.И., и Шмид-Хемпель, П. (1986). Кормление медоносных пчел в центральных местах: влияние времени в пути и потока нектара на наполнение урожая. Behav. Ecol. Sociobiol. 19, 19–24. DOI: 10.1007 / BF00303838
CrossRef Полный текст | Google Scholar
Кац, К., Науг, Д. (2015). Энергетическое государство регулирует компромисс между разведкой и эксплуатацией медоносных пчел. Behav. Ecol. 26, 1045–1050. DOI: 10.1093 / beheco / arv045
CrossRef Полный текст | Google Scholar
Кей, А.(2002). Применение теории оптимального кормодобывания для оценки доступности питательных веществ для муравьев. Экология 83, 1935–1944. DOI: 10.1890 / 0012-9658 (2002) 083 [1935: AOFTTA] 2.0.CO; 2
CrossRef Полный текст | Google Scholar
Келлер И., Флури П. и Имдорф А. (2005). Питание пыльцы и развитие колоний у медоносных пчел — Часть II. Пчелиный мир 86, 27–34. DOI: 10.1080 / 0005772X.2005.11099650
CrossRef Полный текст | Google Scholar
Концманн, С., и Лунау, К. (2014). Различные правила для шмелей, собирающих пыльцу и нектар — лабораторное исследование искусственных цветов, предлагающих разбавленный заменитель нектара и суррогат пыльцы. PLoS ONE 9: e
. DOI: 10.1371 / journal.pone.00
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ли, К. П., Бемер, С. Т., Симпсон, С. Дж., И Раубенхаймер, Д. (2002). Геометрический анализ регуляции питательных веществ у гусеницы широкого профиля Spodoptera littoralis (Boisduval). J. Insect Physiol. 48, 655–665. DOI: 10.1016 / S0022-1910 (02) 00088-4
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Леонхардт, С., Блютген, Н. (2012). Та же, но разная пыльца, собирающая пищу в пчелах и шмелях. Apidologie 43, 449–464. DOI: 10.1007 / s13592-011-0112-y
CrossRef Полный текст | Google Scholar
Лихоро, М., Буль, Дж., Чарльстон, М. А., Меч, Г. А., Раубенхаймер, Д., и Симпсон, С.J. (2014). Моделирование питания на всех уровнях организации, от отдельных людей до суперорганизмов. J. Физиология насекомых . 69, 2–11. DOI: 10.1016 / j.jinsphys.2014.03.004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лихоро, М., Читтка, Л., Ле Комбер, С. К., и Рейн, Н. Э. (2012). Пчелы не используют правила ближайшего соседа для оптимизации маршрутов с несколькими местоположениями. Biol. Lett. 8, 13–16. DOI: 10.1098 / RSBL.2011.0661
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Минахан, Д.Ф. и Брюне Дж. (2018). Сильные межвидовые различия в активности кормления, наблюдаемые между медоносными пчелами и шмелями с использованием миниатюрной радиочастотной идентификации (RFID). Фронт. Ecol. Evol . 6: 156. DOI: 10.3389 / fevo.2018.00156
CrossRef Полный текст | Google Scholar
Николсон, С. В. (2011). Пчелиный корм: химический состав и пищевая ценность нектара, пыльцы и их смесей. Afr. Zool. 46, 197–204. DOI: 10.1080 / 15627020.2011.11407495
CrossRef Полный текст | Google Scholar
Нуньес, Дж.А. (1966). Количественные Beziehungen zwischen den Eigenschaften von Futterquellen und Verhalten von Sammelbienen. Z. Vergl. Physiol . 53, 142–164.
Google Scholar
Пейдж, Р. Э. младший, и Фондрк, М. К. (1995). Влияние отбора на уровне колонии на социальную организацию колоний медоносных пчел ( Apis mellifera L .): Компоненты накопления пыльцы на уровне колонии. Behav. Ecol. Sociobiol. 36, 135–144. DOI: 10.1007 / BF00170718
CrossRef Полный текст | Google Scholar
Панкив, Т.(2003). Изменение направления поведения при поиске пищи у эволюционировавших медоносных пчел тропических и умеренных широт ( Apis mellifera L .). Behav. Ecol. Sociobiol. 54, 458–464. DOI: 10.1007 / s00265-003-0640-1
CrossRef Полный текст | Google Scholar
Панкив Т. и Пейдж Р. Э. младший (1999). Влияние генотипа, возраста, пола и касты на пороги реакции на сахарозу и поведение медоносных пчел в поисках пищи ( Apis mellifera L .). J. Comp. Physiol.А 185, 207–213. DOI: 10.1007 / s0035
379
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Панкив Т. и Пейдж Р. Э. младший (2000). Пороги отклика на сахарозу предсказывают разделение труда у медоносных пчел при кормлении. Behav. Ecol. Sociobiol. 47, 265–267. DOI: 10.1007 / s002650050664
CrossRef Полный текст | Google Scholar
Паоли П. П., Донли Д., Стейблер Д., Сасендранат А., Николсон С. В., Симпсон С. Дж. И др. (2014a).Пищевой баланс незаменимых аминокислот и углеводов взрослой рабочей пчелы зависит от возраста. Аминокислоты 46, 1449–1458. DOI: 10.1007 / s00726-014-1706-2
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Паоли П. П., Уэйклинг Л. А., Райт Г. А. и Форд Д. (2014b). Доля незаменимых аминокислот и Sir2 в рационе влияет на продолжительность жизни пчел. Возраст 36: 9649. DOI: 10.1007 / s11357-014-9649-9
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Пирс, Г.Дж. И Олласон Дж. Г. (1987). Восемь причин, почему теория оптимального кормодобывания — пустая трата времени. Oikos 49, 111–118. DOI: 10.2307 / 3565560
CrossRef Полный текст | Google Scholar
Пирк, К. В. У., Буду, К., Хьюман, Х. и Николсон, С. (2010). Важность типа белка и соотношения белков и углеводов для выживания и активации яичников пчел, содержащихся в клетках ( Apis mellifera scutellata ). Apidologie 41, 62–72. DOI: 10.1051 / apido / 2009055
CrossRef Полный текст | Google Scholar
Пайк, Г.Х. (1984). Теория оптимального кормодобывания: критический обзор. Annu. Rev. Ecol. Syst. 15, 523–575. DOI: 10.1146 / annurev.es.15.110184.002515
CrossRef Полный текст | Google Scholar
Раубенхаймер, Д., Симпсон, С. Дж., И Майнц, Д. (2009). Питание, экология и экология питания: к интегрированной структуре. Funct. Ecol. 23, 4–16. DOI: 10.1111 / j.1365-2435.2009.01522.x
CrossRef Полный текст | Google Scholar
Рулстон, Т.Х. и Кейн Дж. Х. (2000). Пищевая ценность и усвояемость пыльцы для животных. Plant Syst. Evol. 222, 187–209. DOI: 10.1007 / 978-3-7091-6306-1_10
CrossRef Полный текст | Google Scholar
Рюденауэр, Ф. А., Спете, Дж., И Леонхард, С. Д. (2015). Как узнать, какая пища вам подходит: шмели используют вкус, чтобы различать различные концентрации пищи, различающиеся по содержанию питательных веществ. J. Exp. Биол . 218, 2233–2240. DOI: 10.1242 / jeb.118554
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шайнер, Р., Пейдж, Р. Э. младший, и Эрбер, Дж. (2001). Влияние генотипа, кормовой роли и чувствительности сахарозы на тактильное обучение медоносных пчел ( Apis mellifera ). Neurobiol. Учиться. Mem. 76, 138–150. DOI: 10.1006 / NLME.2000.3996
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Schmid-Hempel, P. (1987). Эффективный сбор нектара пчелами I.Экономические модели. J. Anim. Ecol. 56, 209–218. DOI: 10.2307 / 4810
CrossRef Полный текст | Google Scholar
Шмид-Хемпель П., Касельник А. и Хьюстон А. И. (1985). Медоносные пчелы максимизируют эффективность, не наполняя свой урожай. Behav. Ecol. Sociobiol. 17, 61–66. DOI: 10.1007 / BF00299430
CrossRef Полный текст | Google Scholar
Шульц Д. Дж., Хуанг З. Ю. и Робинсон Г. Э. (1998). Влияние нехватки корма в колонии на развитие поведения медоносных пчел. Behav. Ecol. Sociobiol. 42, 295–303. DOI: 10.1007 / s002650050442
CrossRef Полный текст | Google Scholar
Сили Т. Д. (1995). Мудрость улья: социальная физиология колоний медоносных пчел . Кембридж, Массачусетс: Издательство Гарвардского университета.
Google Scholar
Sih, A., и Christensen, B. (2001). Теория оптимальной диеты: когда она работает, а когда и почему не работает? Anim. Behav. 61, 379–390. DOI: 10.1006 / anbe.2000.1592
CrossRef Полный текст | Google Scholar
Сперанца А., Кальцони Г. Л. и Пачини Э. (1997). Наличие моно- или дисахаридов и запасов полисахаридов в зрелых пыльцевых зернах. Секс. Завод Репрод. 10, 110–115. DOI: 10.1007 / s004970050076
CrossRef Полный текст | Google Scholar
Стейблер Д., Паоли П. П., Николсон С. В. и Райт Г. А. (2015). Баланс питательных веществ взрослого рабочего шмеля ( Bombus terrestris ) зависит от пищевого источника незаменимых аминокислот. J. Exp. Биол. 218, 793–802. DOI: 10.1242 / jeb.114249
PubMed Аннотация | CrossRef Полный текст | Google Scholar
vanEngelsdorp, D., Evans, J. D., Donovall, L., Mullin, C., Frazier, M., Frazier, J., et al. (2009). «Зараженная пыльца»: новое состояние в семьях медоносных пчел, связанное с повышенным риском гибели семьи. J. Invertebr. Патол. 101, 147–149. DOI: 10.1016 / j.jip.2009.03.008
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Варжу, Д.и Нуньес Дж. (1991). Что оптимизируют сбор медоносных пчел? J. Comp. Physiol. А 169, 729–736. DOI: 10.1007 / BF00194901
CrossRef Полный текст | Google Scholar
Ваудо А. Д., Патч Х. М., Мортенсен Д. А., Грозингер К. М. и Тукер Дж. Ф. (2014). Шмели проявляют повседневные поведенческие паттерны при поиске пыльцы. Членистоногие. Взаимодействие с растениями. 8, 273–283. DOI: 10.1007 / s11829-014-9312-5
CrossRef Полный текст | Google Scholar
Уэллс, Х.и Уэллс П. Х. (1986). Оптимальный рацион, минимальная неопределенность и индивидуальное постоянство в добыче медоносных пчел, Apis mellifera . J. Anim. Ecol. 55, 881–891. DOI: 10.2307 / 4422
CrossRef Полный текст | Google Scholar
Уилсон, Э. О. (1971). Общества насекомых . Кембридж, Массачусетс: Издательство Гарвардского университета.
Google Scholar
Уинстон, М. Л. (1991). Биология медоносной пчелы . Кембридж, Массачусетс: Издательство Гарвардского университета.
Google Scholar
Нутригеномика у медоносных пчел: цифровой анализ экспрессии генов питательного воздействия пыльцы на здоровых пчел и пчел, зараженных варроа | BMC Genomics
Статистика секвенирования тегов
Четыре библиотеки DGE-тегов были созданы из наших экспериментальных групп: контрольные пчелы без пыльцевого рациона (VP-), контрольные пчелы, которых кормили пыльцой (V-P +), пчелы, зараженные варроа, без пыльцевый рацион (V + P-) и пчелы, паразитирующие на варроа, которых кормили пыльцой (V + P +). Всего было секвенировано около 14 × 10 6 тегов (за исключением последовательностей адаптеров) из каждой библиотеки (подробности см. В Таблице 1).Библиотеки пчел, зараженных варроа, содержали больше выровненных тегов (~ 7,3 × 10 6 тегов), чем непаразитированные пчелы (~ 4,5 × 10 6 тегов). Это большее количество выровненных тегов связано с увеличением числа тегов, соответствующих вирусным последовательностям, которые достигли 1, 417, 481 в библиотеке VP-, 824, 212 в V-P +, 4, 258, 724 в V + P- и 5. , 061, 614 в V + P +. Эти вирусные теги составляют 32% от общего числа выровненных тегов в библиотеке V-P-, 18% в V-P +, 60% в V + P- и 67% в V + P +. Это ясно показывает, что варроа вызывает увеличение количества патогенов у пчел.Вероятно, это было вызвано повторным питанием варроа гемолимфой пчел, что не только влияет на физиологию пчел, но и способствует передаче различных вирусов [19, 21, 27, 28]. Кроме того, мы заметили, что здоровые (не зараженные паразитами) пчелы, которых кормили пыльцой, имели в целом более низкую нагрузку метками патогенов, чем пчелы, ограниченные сахарной диетой. Однако небольшое количество вирусов присутствовало на значительно разных уровнях в каждой группе лечения (см. Влияние кормления пыльцой и паразитизма варроа на распространенность вируса ).
Таблица 1 Статистика секвенирования DGEЭти отдельные теги и их геномный счет, а также необработанные данные доступны из базы данных NCBI Gene Expression Omnibus (GEO) с регистрационным номером: GSE25161. Количество отдельных выровненных тегов было одинаковым для каждой библиотеки (таблица 1), и их распределение показывает согласованный образец для 4 библиотек DGE, при этом большинство отдельных тегов (от 34 до 40%) появляются только один раз, а около 40% — между двумя. и десять раз (дополнительный файл 1).Процент меток с большим числом счетчиков резко упал, и только 1% отдельных меток имеют счетчик выше 1 000. Подобный образец профилирования DGE-тегов ранее наблюдался у голов медоносных пчел [29] и у других насекомых и позвоночных [ 30, 31]. В общей сложности около 8000 генов медоносных пчел были картами из четырех брюшных библиотек (Таблица 1), что составляет примерно 60% генов, идентифицированных на данный момент из генома медоносной пчелы [8]. Для сравнения было обнаружено, что от 55 до 60% генов экспрессируются в мозге пчелы с помощью DGE и анализа микрочипов [29, 32].
Дифференциально экспрессируемые гены и профили экспрессии маркерных генов
Число генов, на уровень экспрессии которых значительно повлияло кормление пыльцой или паразитизм варроа при скорректированном значении P для множественного сравнения (значение Q ) < 0,01 приведено в Таблице 2, а списки представлены в Дополнительном файле 2. В то время как аналогичное количество генов было увеличено и уменьшено питанием пыльцой у здоровых пчел, паразитизм варроа оказал сильное ингибирующее действие на экспрессию генов.Это ингибирование на уровне транскрипции может объяснить потерю веса и снижение метаболизма, вызванное варроа у появляющихся пчел [20].
Таблица 2 Гены, дифференциально транскрибируемые после кормления пыльцой или паразитизма варроаЧтобы оценить надежность нашего профилирования DGE-tag, мы проверили, согласуется ли регуляция некоторых генов-кандидатов с паттерном экспрессии, о котором ранее сообщалось в более ранних исследованиях, и выполнили обратное -транскриптаза ПЦР (ОТ-ПЦР) на эти гены.Вителлогенин (Vg) — это белок желтка, который поглощается развивающимися ооцитами и связан с яйценоскостью у маток [33, 34], но также обладает антиоксидантными функциями, которые защищают пчел от окислительного стресса и увеличивают продолжительность жизни [35, 36]. Следовательно, уровень Vg можно приравнять к показателю долголетия. Поскольку питание пыльцой способствует развитию жирового тела [12, 17], а Vg синтезируется в жировом теле [35], мы ожидали увеличения уровня Vg у пчел, которых кормили пыльцой, как было обнаружено ранее [37].Это было подтверждено анализом DGE и RT-PCR (дополнительный файл 2 и рисунок 1). Это говорит о том, что одним из способов увеличения продолжительности жизни при питании пыльцой [7] является повышение уровня Vg . Интересно, что паразитизм варроа снизил уровень Vg у пчел, которых кормили пыльцой (см. Также [38]), но этот уровень все еще был значительно выше, чем у паразитированных пчел, ограниченных сахарной диетой.
Рисунок 1Гены-маркеры, пораженные питанием пыльцой или паразитизмом варроа .Значения экспрессии 4 генов, выбранных из набора генов, дифференцированно транскрибируемых при разных обработках. Показаны данные ОТ-ПЦР, нормированные на β-актин . Средние значения ± SE показаны для 8 групп по 3 пчелы на обработку. Разные буквы указывают на существенные различия, обнаруженные U-критериями Манна-Уитни.
Другой интересный ген — malvolio ( mvl ), который кодирует трансмембранный переносчик марганца, участвующий в чувствительности к сахарозе [39].У медоносных пчел более чувствительные к сахарозе рабочие-собиратели имеют более высокий уровень мРНК mvl в головном мозге, чем молодые медсестры, ухаживающие за расплодом внутри улья [40]. Пыльца вызвала паттерн экспрессии, противоположный Vg (рис. 1). У пчел, которых кормили только сахаром, в брюшной полости был более высокий уровень мвл. Поскольку старые собиратели питаются в основном углеводами, а молодые медсестры питаются и углеводами, и пыльцой [41], у появляющихся пчел, которые ограничиваются сахарной диетой, как правило, развивается профиль экспрессии mvl , подобный фуражирам, по сравнению с пчелами, которых кормили пыльцой, которая больше похожи на пчел-кормилиц.Действительно, в отличие от плохого питания, диета с высоким содержанием белка может стимулировать выполнение задач медсестры за счет развития гипофарингеальных желез, используемых для выделения личиночной пищи [7]. Точно так же паразитизм v arroa индуцировал повышение уровня mvl (хотя и незначительно у пчел, которых кормили пыльцой), что также указывает на профиль собирателей.
Уровень экспрессии гена пропенолоксидазы ( PPO ), участвующего в иммунитете насекомых [42], также был подтвержден ОТ-ПЦР (рис. 1) и предыдущими данными.Действительно, паразитизм варроа подавлял экспрессию PPO , что согласовывалось с ранее сообщенным иммуносупрессивным действием [21], но кормление пыльцой не влияло на уровень PPO (см. Также [17] на уровне активности фермента).
Наконец, мы проверили уровень экспрессии ортолога Drosophila spaetzle , который является участником иммунного сигнального пути Toll против грибов и бактерий [42]. Профилирование этого гена с помощью DGE-метки, подтвержденное ОТ-ПЦР, показало, что кормление пыльцой значительно увеличивало количество транскриптов spetzle у здоровых пчел (рис. 1), что предполагает, что питательные вещества пыльцы могут усиливать иммунную функцию пчел.Экспрессия этого гена была снижена паразитизмом варроа у пчел, которых кормили пыльцой, снова поддерживая иммуносупрессию, вызванную варроа. В целом, эти результаты подтвердили профилирование DGE-тегов четырех библиотек.
Функциональный анализ дифференциально экспрессируемых генов
Онтологический анализ генов был проведен для изучения того, какие функциональные компоненты регулируются питанием пыльцой и на которые влияет паразитизм варроа. Хотя стенка пыльцы, состоящая из спорополленина и целлюлозы, плохо переваривается пищеварительными ферментами медоносной пчелы, от 50 до 98% содержания пыльцы может быть извлечено из проглоченной пыльцы, в зависимости от типа пыльцы и возраста пчелы [10].Пчелы 9-дневного возраста, близкого к возрасту проанализированных нами пчел, могут переваривать до 95% содержания пыльцы [41]. Повышенная регуляция многих метаболических функций у пчел, питающихся пыльцой, подчеркивает высокую усвояемость пыльцы (рис. 2А). Действительно, в ответ на питательные вещества пыльцы, состоящие в основном из белков и липидов, активируются гены, участвующие в протеолизе, метаболизме липидов и кодирующие пептидазы. Углеводный обмен также активизировался при кормлении пыльцой, что свидетельствует о пищеварительной реакции на крахмал.Помимо метаболических категорий, «окислительное фосфорилирование» и «образование метаболитов-предшественников и энергии» показали значительное обогащение среди тестируемых категорий. В этом нет ничего удивительного, поскольку питательные вещества используются для производства энергии, например АТФ, посредством окислительного фосфорилирования.
Рисунок 2Функциональный (генная онтология) анализ генов, на которые влияет питание пыльцой (A, B) или паразитизм варроа (C, D) . Списки генов, регулируемых питанием пыльцой или паразитизмом варроа, были проанализированы для статистического обогащения ассоциированных терминов GO ( P <0.05) относительно представления этих терминов для всех генов, проанализированных с помощью DGE. Показаны молекулярные функции и биологические процессы, которые были обогащены наборами генов с повышенной (A, C) и пониженной регуляцией (B, D).
Группы генов, участвующих в процессах трансляции и биогенезе рибосом, подавлялись питанием пыльцой (рис. 2В). У всех эукариот биогенез рибосом и регуляция трансляции необходимы для роста тканей и регулируются приемом пищи. Правильная регуляция этих процессов необходима для обеспечения гомеостаза клеток и тканей при изменении питания и подавляется во время голодания по питательным веществам [2].Следовательно, при питании пыльцой ожидалось усиление биогенеза и трансляции рибосом. Однако большая часть потребления пыльцы и развития тканей (например, гипофарингеальных желез, яичников и жирового тела) происходит в течение первой недели после появления взрослых особей [43]. Предыдущее завершение физиологического развития до нашего анализа могло объяснить подавление трансляционных процессов и биогенеза рибосом.
Влияние питания пыльцой на физиологию медоносных пчел резко изменилось паразитизмом варроа.В то время как группы генов, участвующих в биогенезе и трансляции рибосом, также были подавлены, метаболические категории больше не были активированы (рис. 2А). Только небольшие наборы генов, участвующих в протеолизе, активности пептидазы и метаболизме углеводов, все еще активировались питанием пыльцой. Напротив, большинство генов, участвующих в метаболизме белков, подавлялись у пчел, зараженных варроа (рис. 2В), что демонстрирует, что клещи варроа ингибируют переваривание и / или использование белков.Кроме того, категории «липидный метаболический процесс», «окислительное фосфорилирование» и «образование метаболитов-предшественников и энергия» больше не обогащались. Также было обнаружено, что метаболические процессы в основном подавляются паразитизмом варроа в предкуколках [22]. В целом, эти результаты указывают на то, что паразитированные пчелы не могут правильно ассимилировать и использовать питательные вещества пыльцы, необходимые для их физиологического развития.
Это было дополнительно подтверждено действием варроа на транскриптом пчелы.Функциональный анализ показал, что все предыдущие категории, такие как биогенез рибосом, трансляция, метаболические функции и выработка энергии, подавлялись клещом варроа (рис. 2D). Ингибирование варроа биогенеза и трансляции рибосом, необходимых для роста клеток, может приводить к замедлению развития тканей. Более низкие концентрации белков и углеводов, вызванные варроа [20], могут быть объяснены нашими результатами по ингибированию метаболических генов и генов, кодирующих эндопептидазу.Об этом сокращении метаболических резервов, вызванном варроа, сообщалось при появлении [20], поэтому подавление метаболических генов, продолжающееся 8 дней после вылупления у пчел, которых кормили пыльцой, показывает, что питательные вещества пыльцы не могут компенсировать эти потери.
Большой набор генов регулировался кормлением пыльцой как у паразитированных, так и у непаразитированных пчел, и имел сходный паттерн экспрессии (таблица 3, часть 1). Однако подмножество генов активируется при кормлении пыльцой у здоровых пчел и снижается у паразитированных пчел.Аналогичная картина наблюдалась в отношении воздействия варроа на пчел, которых кормили с пыльцой или без нее (таблица 3, часть 2). Чтобы расшифровать биологический смысл этих противоположных правил, мы провели функциональный анализ этих подмножеств. Среди подмножеств, которые регулируются кормлением пыльцой у здоровых пчел, но подавляются у паразитированных пчел, группы генов участвуют в биогенезе рибосом, трансляции, метаболизме белков и выработке энергии (дополнительный файл 3, часть A). Подгруппа, активированная паразитизмом варроа у пчел, питающихся без пыльцы, но подавленная у пчел, питающихся пыльцой, показала значительное обогащение генов, кодирующих оксидоредуктазу и участвующих в метаболизме аминокислот (дополнительный файл 3, часть B).Эти результаты подтвердили отрицательное влияние варроа на те функции, о которых ранее сообщалось. Действительно, энергетический обмен и потребность в питательных веществах у клещей высоки, они используют до 25% пищевых запасов куколок [44]. Уменьшение метаболических резервов у взрослых может быть связано с развитием вирусов, передаваемых клещами на стадии личинок или куколок.
Таблица 3 Перекрытие генов, регулируемых в брюшной полости питанием пыльцой или паразитизмом варроаРезультаты, аналогичные функциональному анализу, были получены при анализе путей (дополнительный файл 4).Используя более поздний анализ, мы также изучили влияние пыльцы и варроа на аминокислоты. Действительно, медоносным пчелам для развития взрослых требуется десять незаменимых аминокислот: аргинин, гистидин, лизин, триптофан, фенилаланин, метионин, треонин, лейцин, изолейцин и валин [45]. Это означает, что эти аминокислоты не могут быть синтезированы пчелами de novo и поэтому должны поступать непосредственно в рацион либо в виде свободных аминокислот, либо перевариваться из белков. Поэтому мы изучили пути метаболизма аминокислот, которые были значительно обогащены после кормления пыльцой, и обнаружили, что четыре из них (аргинин, лейцин, изолейцин и валин) были активированы (дополнительный файл 4).Интересно, что пять из этих аминокислотных путей подавляются варроа (лизин, триптофан, лейцин, изолейцин и валин), подтверждая, что варроа значительно снижает развитие пчел. Однако ни один из этих незаменимых аминокислотных путей не был существенно затронут у пчел, зараженных варроа, которых кормили пыльцой, по сравнению с паразитированными пчелами, ограниченными сахарной диетой, что позволяет предположить, что питательные вещества пыльцы компенсируют негативное влияние варроа на метаболизм этих аминокислот.
Влияние кормления пыльцой и паразитизма варроа на путь восприятия питательных веществ
Путь инсулина / TOR представляет собой консервативный сигнальный каскад, который функционирует как путь восприятия питательных веществ, связывая потребление пищи с ростом и метаболизмом животных, включая размножение и продолжительность жизни [46 , 47]. У медоносных пчел этот путь играет важную роль в регуляции старения особей [35, 48], но также и на социальном уровне, регулируя разделение труда между рабочими пчелами [49, 50].Меньше известно о регуляции этого пути у медоносных пчел. Однако недавнее исследование показало, что ингибирование экспрессии субстрата рецептора инсулина в периферических тканях (в отличие от центральных тканей мозга) заставляло пчел-собирателей собирать пыльцу, а не нектар [51]. Анализируя профилирование DGE-tag периферических тканей (брюшной полости), мы обнаружили, что большинство генов сигнальных путей mTOR активировалось питанием пыльцой у здоровых пчел (рис. 3). Экспрессия инсулиноподобного пептида 2 ( ILP-2 ) заметно увеличивалась в ответ на питание пыльцой, как было обнаружено в ответ на аминокислоты [48], что может способствовать физиологии медсестры, которая продлевает продолжительность жизни пчел [52].В совокупности эти результаты показывают, что путь инсулина / TOR положительно реагирует на питание пыльцой и может запускать индуцированное пыльцой физиологическое развитие. Противоположный эффект наблюдался в отношении ILP-1 , с более высокой экспрессией в плохих условиях питания [50]. Однако эти обратные паттерны могут быть фактически объяснены функциональными различиями ILP-1 и ILP-2 [52]. У пчел, зараженных варроа, питание пыльцой подавляло этот путь.Это может быть вызвано более высокой распространенностью вирусов у паразитированных пчел, которых кормили пыльцой, по сравнению с паразитизированными пчелами, которых кормили только сахаром (см. Влияние кормления пыльцой и паразитизма варроа на распространенность вируса ). Действительно, паразитизм варроа (в сочетании с более высокой вирусной нагрузкой) индуцировал аналогичный паттерн пути TOR (рис. 3). Паразит подавляет метаболизм белков и снижает запасы питательных веществ у пчел (см. Выше), что можно объяснить подавлением пути TOR, опосредующего рост тканей в ответ на потребление питательных веществ.Это дополнительно подтверждается ингибированием slimfast паразитизмом варроа у пчел, питающихся пыльцой. Подавление этого гена, который кодирует переносчик аминокислот и является нижестоящей мишенью пути TOR, вызывает недостаточный глобальный рост у Drosophila , подобный тому, который наблюдается у людей, выросших в плохих условиях питания [46, 53].
Рисунок 3Путь передачи сигналов mTOR, пораженный питанием пыльцой или паразитизмом варроа .Гены из сигнального пути mTOR дрозофилы , которые дифференциально транскрибировались после кормления пыльцой или паразитизма варроа. Путь был изменен из KEGG: Kyoto Encyclopedia of Genes and Genomes http://www.kegg.com/. Серый цвет указывает на наличие генов в геноме пчелы. Квадраты и кружки указывают на гены и расширенные функции, которые были активированы и подавлены между двумя видами лечения, соответственно.
TOR, как известно, реагирует на присутствие аминокислот и вызывает активацию биогенеза рибосом, трансляции [2, 54–57] и энергетического метаболизма [58, 59], необходимых для роста тканей.Интересно, что за подавлением пути TOR следовало подавление каждого биологического процесса (рис. 3), что указывает на то, что те же механизмы пути восприятия питательных веществ обнаруживаются у медоносных пчел. Активация этого пути у пчел, получавших пыльцу, сопровождалась увеличением энергетического метаболизма, но биогенез и трансляция рибосом подавлялись. Как объяснялось выше, возможно, что развитие ткани, требующее активации трансляционных процессов, уже завершилось.
Уровень экспрессии генов не всегда хорошо соответствовал функциям активатора / ингибитора, установленным в пути Drosophila TOR. Паттерны экспрессии соответствуют глобальным профилям DGE-tag целого брюшка пчелы, но многие гены обладают тканеспецифичными функциями. Следовательно, нацеливание на определенные ткани, такие как жировое тело, которое выполняет важные функции хранения, связанные с питанием, должно обеспечить более точные данные экспрессии для анализа путей.
Влияние кормления пыльцой и паразитизма варроа на гены, влияющие на продолжительность жизни и гены иммунитета
Поскольку питание пыльцой и варроа увеличивает и сокращает продолжительность жизни пчел [12, 60] соответственно, мы рассмотрели паттерн экспрессии обнаруженных генов. чтобы повлиять на продолжительность жизни Drosophila .Эти гены были обнаружены с помощью функционального анализа из биоинформатических ресурсов DAVID 6.7 [61] и GoToolbox [62]. Мы обнаружили, что 20 генов, в том числе Vg , которые, как известно, влияют на продолжительность жизни, по-разному экспрессируются при разных видах лечения. Мутация в локусах каждого из этих генов вызывает снижение продолжительности жизни, за исключением Tor , ингибирование которого увеличивает продолжительность жизни [63]. Мы ожидали найти среди этих генов больший набор, который будет активироваться при питании пыльцой, чем снижаться, и наоборот, в отношении эффекта варроа.Согласно нашим ожиданиям, 7 из 10 генов, регулируемых потреблением пыльцы, были активированы у непаразитизированных пчел (рис. 4A). Например, Sod и Trxr-1 , которые кодируют цитоплазматическую Cu-Zn-супероксиддисмутазу и тиоредоксинредуктазу, соответственно [64], как известно, увеличивают продолжительность жизни [65, 66] благодаря разложению супероксидных радикалов. и H 2 O 2 или нейтрализация активных форм кислорода. Положительный эффект кормления пыльцой на характер экспрессии обоих генов был подтвержден анализом ПЦР с обратной транскриптазой (ОТ-ПЦР), выполненным на другой группе экспериментальных пчел (n = 8 пулов пчел на обработку, дополнительный файл 5).Эти гены могут быть вовлечены в вызванные пыльцой молекулярные механизмы, которые увеличивают продолжительность жизни медоносных пчел. Однако Sod 2 (митохондриальная супероксиддисмутаза Mn), которая также участвует в деградации супероксидных радикалов и H 2 O 2 [64], не подвергалась воздействию пыльцы (дополнительный файл 5), в отличие от подавление, обнаруженное с помощью анализа DGE. Из-за более высокого уровня Vg (Рисунок 1) ожидается более продолжительная продолжительность жизни паразитированных пчел, получающих пыльцу, по сравнению с паразитированными пчелами, которых кормили только сахаром (Рисунок 4A).Однако, за исключением Vg , все гены, которые регулировались потреблением пыльцы, были подавлены. Может ли питание пыльцой влиять на устойчивость к варроа, еще предстоит проверить, но возможно, что влияние пыльцы на гены, влияющие на продолжительность жизни, будет компенсировано увеличением Vg. Картина была более ясной в отношении влияния варроа на 19 генов, влияющих на продолжительность жизни (рис. 4A и дополнительный файл 5). Все они были подавлены (в том числе Sod 2, в присутствии пыльцы, дополнительный файл 5), что свидетельствует об уменьшении продолжительности жизни, вызванном варроа.
Рисунок 4Иммунные гены и гены продолжительности жизни, пораженные питанием пыльцой или паразитизмом варроа . A) Гены из наших наборов генов, которые участвуют в продолжительности жизни у Drosophila и медоносных пчел (т.е. Vg ). Б) Иммунные гены, обнаруженные у медоносных пчел (список генов взят из [42]). Цветовая шкала на тепловой карте (от зеленого к красному) показывает коэффициенты транскрипции log2. Для каждого гена указывается инвентарный номер и аннотация.
Затем мы исследовали влияние потребления пыльцы и варроа на характер экспрессии иммунных генов, которые были ранее аннотированы [42].Некоторые иммунные гены активируются при кормлении пыльцой здоровых пчел (рис. 4В). Три из них кодируют антимикробные пептиды (AMP) ( Lyzozyme-2 и -3, Defensin-1 ), что может быть связано с более высоким содержанием жирового тела, основного сайта синтеза AMP. Интересно, что в отсутствие иммунного заражения (здоровые пчелы) Spaetzle и ген, кодирующий белок распознавания пептидогликана ( PGRP ) — LC , которые являются активаторами путей Toll и Imd соответственно, оба были усилены кормлением протеином.Повышающая регуляция этих ключевых иммунных генов, подтвержденная анализом ОТ-ПЦР (Рисунок 1 для Spaetzle и Дополнительный файл 5 для Defensin-1 и PGRP — LC ), предполагает, что пчелы могут иметь эффективную иммунную систему. ответ на патогенные инфекции. У паразитированных пчел только Imd , специфический ген антимикробной защиты, активировался при кормлении пыльцой. Небольшое увеличение экспрессии Imd также наблюдалось при анализе RT-PCR, хотя и не значимо (p = 0.058, доп. Файл 5). У здоровых пчел экспрессия Imd не изменилась после кормления пыльцой (дополнительный файл 5). Поскольку белковое и аминокислотное питание может усиливать иммунные функции у людей [4, 67] и насекомых [68, 69], ожидалось повышение активности иммунных генов. Функциональный анализ показал, что у зараженных варроа пчел, которых кормили пыльцой, подавлялось значительное количество генов, участвующих в метаболизме белков, по сравнению с паразитированными пчелами, которых кормили только сахаром (рис. 2В).Дефицит белкового обмена мог повлиять на иммунную систему этих пчел. Это иммуносупрессия может быть объяснена более высокой вирусной нагрузкой у паразитированных пчел, которых кормили пыльцой, по сравнению с паразитизированными пчелами, которых кормили только сахаром (см. Влияние кормления пыльцой и паразитизма варроа на распространенность вируса ). Однако при кормлении пыльцой паразитированные пчелы могут вкладывать больше энергии в другие защитные механизмы, чем иммунитет, который подавляется варроа, например антиоксидантная система (более высокий уровень Vg у пчел, питающихся пыльцой).
Варроа индуцировал ингибирование большинства иммунных генов у пчел, которых кормили с пыльцой или без нее (включая PGRP — LC , дополнительный файл 5). Однако несколько иммунных генов были активированы, например, AMP ( Defensin-1, Hymenoptaecin, Apidaecin-1 и Lyzozyme-3 ) и cact-1 , который контролирует продукцию AMP [70]. Активная регуляция Defensin-1 паразитами варроа была подтверждена анализом ОТ-ПЦР (дополнительный файл 5).Эти гены, за исключением Lyzozyme-3 , уже были активированы иммунным стимулом [42]. Механизмы, запускающие иммуносупрессию, все еще неправильно поняты, но одно из объяснений может заключаться в том, что недостаточный метаболизм белка, вызванный варроа и ассоциированными вирусами (см. Выше), блокирует развитие иммунной функции.
Иммунные гены, которые были активированы кормлением пыльцой и варроа (рис. 4В), принадлежали к семейству антибактериальных пептидов или были ключевыми активаторами иммунных путей.Поскольку характеристика иммунной защиты медоносной пчелы еще находится в зачаточном состоянии, возможно, что другие иммунные гены на самом деле не играют важной роли в иммунном ответе пчел. В самом деле, гены иммунного сигнального пути обычно играют разные роли, например, функции развития.
Влияние кормления пыльцой и паразитизма варроа на распространенность вируса
Используя библиотеки DGE-tag из образцов брюшной полости медоносных пчел, мы провели метагеномный анализ, чтобы определить влияние кормления пыльцой и варроа на распространенность 10 вирусов медоносных пчел: РНК 1 вируса паралича пчел, РНК 2 вируса хронического паралича пчел, вирус Sacbrood, вирус деформированного крыла, вирус черной маточной клетки, вирус острого паралича пчелы, вирус пчелы Кашмира, вирус Какуго, вирус 1 деструктора Варроа и вирус 1 острого паралича Израиля.Все они были идентифицированы в наших библиотеках DGE-тегов, но очень небольшое количество копий тегов РНК 1 вируса хронического пчелиного паралича, РНК 2 вируса хронического пчелиного паралича, вируса Сакбруд, вируса черной матки, вируса острого паралича пчел, вируса пчел Кашмира и вирус острого паралича Израиля (от 1 до 65, см. данные, депонированные в базе данных GEO: GSE25161), и не было обнаружено различий в их численности между обработками. Однако метки, отображающие вирус деформированного крыла (DWV), вирус Varroa destructor, (VDV) и вирус Какуго (KV), были широко распространены и в значительной степени зависели от различных обработок (таблица 4). Varroa destructor является эффективным переносчиком вирусов пчел, способствуя их передаче и развитию (DWV [19, 21], VDV [27], KV [28]), но присутствие вирусов у контрольных пчел можно объяснить вертикальной передачей от маток и трутней к потомству рабочих через оплодотворенные яйца [71].
Таблица 4 Влияние кормления пыльцой и паразитизма варроа на распространенность вируса медоносных пчелКак и ожидалось, варроа спровоцировал ускоренное развитие вируса (Таблица 4).Поскольку AMP обладают антимикробной активностью широкого спектра против бактерий, дрожжей, грибов, паразитов, а также вирусов [72], активация AMP у пчел, паразитирующих на варроа (рис. 4B), может представлять иммунный ответ на вирусную инфекцию с высоким уровнем вирусной нагрузки. Различные исследования показали, что питательные вещества пыльцы могут помочь пчелам бороться с паразитами и патогенами [14, 15]. Например, недавнее исследование показало, что питание пыльцой снижает распространенность DWV у здоровых взрослых пчел [16]. Мы получили аналогичные результаты на здоровых (непаразитированных) пчелах с пыльцевым питанием, снижающим нагрузку ВДВ.Однако мы наблюдали обратную картину с DWV и KV. Вирусы используют клеточный аппарат для размножения, поэтому большее количество доступных клеток также может способствовать лучшему развитию вирусной популяции. Например, было обнаружено, что DWV размножается в теле пчелиного жира [73], и эта ткань более развита у пчел, питающихся пыльцой. Мы также обнаружили, что кормление пыльцой увеличивает распространенность вирусов у пчел, зараженных варроа, что указывает на то, что питание пыльцой неэффективно для ограничения вирусной нагрузки у сильно инфицированных пчел.Напротив, более крупный физиологический механизм может способствовать их размножению, но он также может помочь пчелам противостоять вирусной инфекции.
Практический пример ведения бизнеса Sparks & Honey
Sparks & Honey (S&H) из Нью-Йорка — это крупномасштабная культурная консалтинговая компания, которая стремится решать глобальные проблемы с помощью социальных наук и наук о данных. Основываясь на убеждении, что люди в одиночку не могут выполнить эту миссию, компания S&H создала систему активного обучения, называемую Q ™, которая объединяет людей и машины для осмысления мира в реальном времени.Корпоративный брифинг по культуре Рабочая группа проводит ежедневные брифинги, лежащие в основе системы культурной разведки S&H, помогая разобраться в тысячах точек данных почти в реальном времени. Культурное понимание, полученное в ходе брифингов, помогло консалтинговой компании достичь 70-процентного годового роста доходов в период с 2014 по 2016 год, поскольку рабочая группа улучшила то, как S&H воспринимает и воспринимает сигналы. 1
Фирма стремится идентифицировать, называть и классифицировать сдвиги в культуре до того, как они станут основными тенденциями, а затем стремится помочь клиентам понять и сформировать их, используя таксономию тенденций.В широком смысле, Sparks & Honey воспринимает мир как неструктурированные данные, смесь быстрой и медленной культуры: от твита, новой семантики, вирусного видео до новой политики, академических исследований, поданных патентов, создания стартапов и сделок с венчурным капиталом. потоки. В своем ежедневном отчете рабочая группа по культуре структурирует данные, маркируя отдельные элементы («сигналы») для таксономии тенденций и применяя методологии оценки при отслеживании тенденций во времени.
Консультационный отдел новостей культуры работает круглосуточно, без выходных, 365 дней в году.Фирма привлекает широкий спектр лидеров мнений и культурных наблюдателей со всего мира 2 и применяет комбинацию патентованных методологий, инструментов (см. Рисунок 1), алгоритмов и человеческого понимания для обеспечения культурного интеллекта, анализа данных и стратегий для широкий спектр организаций и брендов. Организации от McDonald’s до DARPA 3 работают с консультантами, чтобы выявить эти культурные сдвиги и принять меры в связи с ними. 4
«Мы превращаем культуру в возможности для брендов в реальном времени», — говорит генеральный директор Терри Янг.«Некоторые из этих вещей развиваются очень быстрыми темпами, и о некоторых из этих вещей вам следует знать, если вы собираетесь продолжать формировать свою организацию через два-три года». 5
С миссией «открывать сознание и создавать возможности в настоящем, будущем и будущем для брендов» s&h стремится быть фильтром и провозвестником идей, которые влияют на культуру и формируют ее. Комментируя отчет консалтинговой компании за 2015 год о появлении нового языка для описания гендерного спектра, например, The Advocate отметила, что «в ее маркетинговых рекомендациях заложена мыслительная бомба радикализма, способная по-настоящему изменить общество. 6 Другие отчеты об активном использовании марихуаны, влиянии поколения X на средний возраст и аналогичных сдвигах отражают масштаб и амбиции компании.
Рабочая группа: брифинг по культуре
В основе деятельности фирмы находится рабочая группа по брифингу по культуре, которая воплощает в жизнь культурный интеллект консалтинговой компании и держит команды клиентов в курсе последних культурных тенденций. Эта группа соответствует нашим критериям для рабочей группы:
- Размер: Основная группа состоит из семи участников, в том числе двух со-брифингов, двух специалистов по обработке данных, модератора, сетевого соединителя, который маркирует сигналы конкретным консультантам для дополнительных исследований или контекста, а также иллюстратора, который создает визуализации ежедневная производительность рабочей группы. 7
- Устойчивое участие: Члены рабочей группы проводят большую часть каждого рабочего дня, работая вместе, чтобы курировать, представлять и анализировать сигналы, обсуждаемые на ежедневном брифинге.
- Комплексные усилия: Используя специально разработанную фирмой систему активного обучения Q ™, сотрудники собирают данные о тенденциях, разрабатывают исследования, отслеживают определенные «сигналы» и курируют мероприятие по культурному брифингу для всей организации S&H (расположенной в Нью-Йорке, Лос-Анджелес) , Сан-Франциско и Чикаго), а также глобальных скаутов и гостей, включая клиентов, которые присутствуют в прямом эфире или смотрят прямую трансляцию.Группа действует совместно, выполняя большую часть работы по отбору сигналов, составлению презентации и анализу идей брифинга.
Каждый день участники просматривают базу данных «культурных сигналов» — всего, от «быстро распространяющегося твита, песни или мема, до популярных новостных статей, исследовательских работ и файлов патентов, до изменений в государственной политике или новых технологий» 8 — для просмотра дополнений, сделанных за последние 24–48 часов, затем выберите около 30 пунктов для представления на брифинге для всей компании в этот день.Эти сигналы — от тенденций в потреблении белка насекомых до ранних признаков «премиализации воздуха» и шрифта Брайля на одежде до рассказа о том, как собак можно считать дрессированными детьми, — выбираются на основе нескольких факторов. Например, имеет ли конкретный сигнал высокую энергию? Помогает ли это установить закономерность, основанную на других, недавно обсуждавшихся сигналах? Представляет ли он новую концепцию на краю культуры? Вносит или подтверждает культурное напряжение (сосуществование противоположных тенденций)?
Рабочая группа не только курирует сигналы — она стремится представить эти сигналы в привлекательной форме, которая вызывает широкие и продуктивные отклики со стороны участников за пределами рабочей группы.Члены решают, как представить сигналы дня, чтобы они были краткими, но вызывающими воспоминания; визуальные эффекты часто столь же эффективны, как и язык, часто с изображениями, взятыми непосредственно из исходных данных. Цель обоих мероприятий — вовлечь в обсуждение умы более широких кругов консультантов, глобальную сеть разведчиков и консультантов по культуре, а также всех гостей, которые пришли, чтобы распознать культурные сигналы посредством обсуждения.
Консультации полагаются на полдник, чтобы подпитывать свою работу с клиентами. Знакомя участников с идеями, источниками данных и интересами, которые они в противном случае могли бы упустить, брифинг помогает противостоять личным или институциональным предубеждениям.Формат «позволяет нам расширить наш анализ по категориям, времени и пространству, взять несколько источников неструктурированных данных, сразу же добавить структуру, а затем использовать обсуждения в реальном времени и междисциплинарные обсуждения для быстрого развития идей и выявления закономерностей», — говорит главный операционный директор. Пол Батлер. «Брифинг также является форумом для изучения и мониторинга культурной напряженности, ключевой методологии в нашем процессе. Мы наблюдаем, что противоположные тенденции могут проявляться и двигаться в культуре одновременно ». Формат — это способ, с помощью которого консультанты могут собраться вместе, чтобы выявить и распознать различные противоречия.Затем они работают с клиентами, чтобы определить, как, когда и где они могут опереться на ту или иную сторону культурного напряжения.
В этом тематическом исследовании мы рассматриваем практику на двух уровнях: внутри самой брифинговой рабочей группы и среди более чем 50 участников ежедневного брифинга. Поскольку более крупная научно-исследовательская организация полагается на результаты ежедневного брифинга и также участвует в нем, в то время как рабочая группа по брифингу создает ежедневный брифинг и также участвует в нем, некоторые методы используются как внутри рабочей группы (при разработке и подготовке к ежедневному брифингу). брифинг) и в самом брифинге.Хотя нас в первую очередь интересует, что происходит внутри самой рабочей группы, в данном случае это не всегда четкое различие — и некоторые практики более заметны в контексте часового брифинга, даже если рабочая группа также использует их для выполнения это работает.
Результаты
Как результат рабочей группы, insights может показаться довольно туманным. Но качество, своевременность, глубина, широта и культурная значимость той информации, которая предоставляется клиентам — в сочетании с интеллектуальным лидерством, анализом и стратегиями — это то, что приносит доход организации.Ежедневный брифинг по культуре — как посредством генерируемых идей, так и путем обучения алгоритмов и персонала распознаванию закономерностей и выявлению тенденций — является ключевым фактором, позволяющим компании S&H выполнять ту работу, которую ценят ее клиенты. За последние три года выручка консалтинговой компании увеличилась на 70% по сравнению с аналогичным периодом прошлого года, и руководители отмечают эффективные ежедневные брифинги за способность консультантов предоставлять дифференцированную информацию, которая способствует росту доходов. Со временем рабочая группа по брифингу по культуре разработала показатель — соединений на сигнал — который, по мнению участников, оказался полезным показателем эффективности брифинга.
Число подключений на сигнал
Учитывая, что ежедневный брифинг по культуре является частью более крупной системы разведки в S&H, рабочая группа отслеживает взаимосвязь, важность и актуальность между сигналами, обсуждаемыми в брифинге, и другими частями системы. Например, связан ли конкретный сигнал с другими тенденциями? Имеет ли это отношение к работе членов нашего консультативного совета или других участников человеческого сообщества? Этот показатель помогает рабочей группе оценить, эффективно ли брифинги расширяют кругозор участников, чтобы лучше понять человеческое поведение и ценности в более широком контексте и информировать краткосрочные кампании и долгосрочные стратегии.Среднее количество и разнообразие связей, установленных между темами, постоянно отслеживаются, чтобы поддерживать то, что группа считает правильным балансом между количеством и качеством для оптимальной производительности — в идеале от трех до пяти соединений на сигнал, обсуждаемых во время брифинга, при этом охватывая широкий спектр сигналов .
Рабочая группа является стержнем стратегии организации, потому что более качественные брифинги приводят к более обширным и важным выводам для клиентов. В ходе успешного брифинга группа могла бы проводить сравнения глобальных рынков как индикаторы изменений в поведении потребителей и человеческих ценностях и искать другие индикаторы того, как эти сдвиги могут повлиять на бизнес и бренды в разных категориях.Например, когда автоматизированная сенсорная система добавила сигнал о разнице в продажах роскошных часов в США и Китае, это привело к отслеживанию более широкой тенденции в восприятии роскоши, которая начала расходиться в двух странах.
Игровые практики
Рабочая группа по брифингу по культуре демонстрирует шесть пересекающихся практик: Стремитесь к общему результату, Ищите новые контексты, Максимально увеличивайте потенциал трений, Развивайте трение, Устраняйте непродуктивное трение и Размышляйте больше, чтобы учиться быстрее .
Приверженность общему результату
Члены рабочей группы стремятся каждый день проводить увлекательные, провокационные и продуктивные брифинги, чтобы разобраться в последних культурных сигналах, полученных из самых разных сфер жизни, Интернета и человеческой сети участников со всего мира. Дневные брифинги собирают весь персонал на час каждый день. Как правило, темп быстрый, обсуждение обманчиво дисциплинировано, а обсуждаемые темы могут варьироваться от смешных до серьезных, тривиальных до потрясающих.Брифинг служит пространством для консалтинговой компании в целом, чтобы ощутить (и разобраться в них) широкий спектр, казалось бы, несопоставимых тенденций, но это также своего рода лаборатория, место, где могут возникнуть интересные идеи и инновации.
Брифинги по культуре считаются центральными в том, что Sparks & Honey предлагает клиентам, и, поскольку многие клиенты посещают брифинги, иногда бывает трудно отделить результаты — идеи, содержание и действия — от практик, которые порождают понимание.Практика рабочей группы предназначена для того, чтобы все лучше и лучше выявлять культурные сигналы, которые можно использовать для прогнозирования и выработки действенных идей и стратегий.
Частью стремления группы к увлекательному, эффективному и продуктивному брифингу является отработка всех сигналов, которые они выбрали для выделения в ходе одночасового занятия. Помимо поддержания определенного количества сигналов, они также рассматривают их влияние на результат как комбинацию факторов:
Открытие нового: Раскрыли ли мы и расширили ли мы себя и наших клиентов новыми идеями, которые могут привести нас на путь трансформации?
Обсуждение и открытие грани : Мы считаем, что то, что происходит на грани культуры, имеет значение и может быть тем, в чем может развиваться будущее бизнеса или категории.
Искать новые контексты
Весь персонал S&H, а также обширная глобальная сеть консультантов и разведчиков, которые перемещаются в различных контекстах, постоянно регистрируют и маркируют культурные сигналы в частной базе данных, которая является частью Q ™, когда они сталкиваются с ними. Автоматизированные персонажи — вне зависимости от пола, расы, политических взглядов, этнической принадлежности, интересов, занятости, образования, возраста, состава домохозяйства и географического положения — также помечают предметы, чтобы обеспечить дополнительную линзу и предвзятое отношение к культуре.Рабочая группа по брифингу каждый день рассматривает весь материал, который был зарегистрирован или помечен в базе данных, как появляющийся тренд или культурный сигнал, подбирая его для ежедневного брифинга. Хотя у группы нет установленной цели по количеству культурных сигналов, которые нужно маркировать каждый день, группа постоянно стремится включать и анализировать сигналы, полученные из различных контекстов, чтобы оставаться на связи с границами культурных изменений, а также помочь понять и подтвердить тенденции.
Например, рабочая группа может намеренно выделить историю о дроне на Среднем Западе, связав его с тем, как меняется городское воздушное пространство.Группа, иногда с клиентами, будет проводить культурных походов — живое исследование культурного опыта и событий — как способ обмена и сбора информации. В другом примере поиска контекста сокобриферы посетили школу ди-джеев в Нью-Йорке, чтобы узнать о курировании и презентации профессиональных ди-джеев. Рабочая группа признала уникальные требования формата брифинга, уравновешивая открытую дискуссию на форуме, которая требует структуры и порядка, чтобы быть эффективной, и увидела прямые параллели между этим и работой ди-джея — объединение отдельных, казалось бы, несвязанных элементов в единое целое. в течение фиксированного периода времени; управление энергией разнообразной толпы известных и неизвестных; презентационные и перформативные аспекты; и потребность поддерживать и достигать определенной частоты вращения педалей, чтобы оказать влияние.
Для высоко концептуальной работы есть еще один важный практический компонент. Оказывается, отслеживания и наблюдения за тенденциями из-за экрана зачастую недостаточно. Когда один из клиентов хотел узнать больше о потреблении верблюжьего молока, рабочая группа по брифингу потратила время на то, чтобы выпить его, углубившись в контекст и получив собственную неприукрашенную правду, поскольку, как согласились участники, блоги и фотографии в Instagram не могли передать уникальный опыт употребления спиртного. молоко. Члены также рассмотрели различные виды «молока», которые были популярны в Соединенных Штатах и других культурах на протяжении многих лет, как долго длилась каждая тенденция и какое молоко прочно закрепилось на рынке.
Максимизировать потенциал трения
Хотя основные роли в рабочей группе по брифингу относительно стабильны, другие целенаправленно меняются. Два-три сотрудника, каждый из которых представляет разных клиентов S&H, присоединяются к рабочей группе в качестве дополнительных членов на несколько дней. Эти сменяющиеся члены из клиентских команд привносят новую энергию, навыки и перспективы — наряду с признаком конкретного клиента — которые могут изменить то, как рабочая группа курирует, интерпретирует и представляет сигналы, а также то, как она готовится и способствует брифинг.Вращающиеся члены вносят некоторый дискомфорт и новые трения в повседневную работу группы. Это может привести к новым подходам к выбору сигналов или новым способам группировки и представления их для получения различных ответов. Например, сменяющийся член, который представляет команду косметических клиентов, может знать, что клиента особенно интересуют тенденции рынка старше 60 лет, и заставлять группу учитывать долголетие при выборе сигналов. Основные члены обеспечивают определенный уровень преемственности, но сменяющиеся члены предлагают им избегать рутины свежими идеями и новыми проблемами клиентов.
Для фактического ежедневного брифинга рабочая группа собирает самых разных участников, включая глобальных скаутов, консультантов, клиентов и других заинтересованных представителей общественности, в рамках концепции «открытого консультирования» s&h. Группа даже начала транслировать в прямом эфире некоторые из своих ежедневных брифингов на Facebook, что еще больше расширило круг участников, хотя она все еще экспериментирует с тем, как лучше всего взаимодействовать с открытой онлайн-аудиторией. Открытое консультирование преследует целенаправленную бизнес-цель: стимулировать продуктивные дебаты и содержательные беседы друг с другом и с внешним миром.Приглашая группу посторонних на брифинг, рабочая группа увеличивает вероятность разногласий и расхождения во взглядах и новых способов интерпретации сигналов. Важно отметить, что это может увеличить количество и глубину связей, которые члены могут устанавливать между сигналами, поскольку неожиданный входной сигнал может обнажить скрытые связи, добавляя реальную глубину и ценность пониманию.
Культивировать трение
На каждом ежедневном сеансе рабочая группа приглашает кого-то из сторонних организаций, чтобы они помогали в качестве помощника.Ротация позволяет другим в организации увидеть, что входит в брифинг, и позволяет рабочей группе по-новому взглянуть на презентацию. Например, один из коллег, называющий себя интровертом, бросил вызов склонности к быстрому, энергичному и непрерывному обмену мнениями, введя преднамеренные периоды молчания. Теперь рабочая группа встраивает тишину в рабочие заседания и брифинги, чтобы побудить людей сосредоточиться на том, что говорится, больше, чем на подготовке к выступлению. Структурирование молчания — встраивание его в подход — привело к тому, что во взаимодействиях меньше внимания уделялось личным реакциям, а больше — сигналам и идеям.Это помогло сместить брифинги с презентаций людей, погруженных в тенденции, в сторону сбора информации и ответов на эти тенденции.
Наконец, рабочая группа берет за правило регулярно менять правила, проводя экспериментальные брифинги, в которых используются новые подходы и средства обмена, в первую пятницу каждого месяца. Встряхивая рутину, эти эксперименты предназначены для того, чтобы вынудить участников выйти из зоны комфорта, что создает трения как в группе, так и в организации.Члены обращают внимание на то, приводит ли трение к пониманию новых или более значимых способов или ведет к более эффективным или продуктивным брифингам. Например, на одном брифинге они использовали тему «Думай как преступник» в качестве эксперимента с целью выработать понимание, выходящее за рамки норм «приемлемого» (или законного) поведения и ценностей, как способ расширить мышление группы до разрабатывать новые инновации в области безопасности и конфиденциальности, комбинировать новый контент или другие идеи будущих продуктов. Этот эксперимент застрял, и теперь участники регулярно используют эту тему в качестве основы для оспаривания предвзятых представлений или для инноваций.
Другим примером является введение рабочей группой брифингов специального выпуска , призванных встряхнуть динамику и привнести больше внешних перспектив в среду брифингов. На этих сессиях группа приглашает внешних экспертов из определенной вертикали, а не сотрудников НИОКР, для обсуждения сигналов и тенденций. Эти сеансы приобретают несколько иной характер, поскольку гости менее готовы реагировать на неявные сигналы и принятые практики ежедневных брифингов.
Одно дело — собрать разных участников, другое — выявить разные точки зрения и проблемы этих участников. Один из способов сделать это — поощрять участие скаута или гостя (учитывая, что большинство гостей приглашены кем-то в s&h, который кое-что знает об их биографии), когда они чувствуют, что у кого-то может быть другая точка зрения на проблему. Рабочая группа представила роль модератора на брифингах, отчасти для того, чтобы побудить и стимулировать диалог за столом между всеми участниками.Роль модератора заключалась в том, чтобы задавать вопросы не участникам совещания, а другим членам рабочей группы или непосредственно гостям, имеющим знания или опыт в данной теме. Этот небольшой сдвиг помог переориентировать комнату и добиться более круглого стола.
Несмотря на то, что s&h поддерживает разнообразие, все еще существуют общие предубеждения — например, все члены рабочей группы проживают в Нью-Йорке. Вместо того, чтобы бороться с этими ограничениями, группа методично определяет их, исходя из теории, что воздействие и анализ могут уменьшить связанные с ними слепые зоны.Все участники четко определяют свои собственные области интересов и подверженности и стараются не скрывать, как они влияют на их внимание и объективы. Ожидается, что от каждого человека добавится уникальный талант восприятия. Рабочая группа использует брифинг как инструмент, чтобы постоянно выявлять и исследовать свои собственные слепые зоны и ломать собственные алгоритмы для поддержки более широкого исследования и участия в культуре. Открытое и непредвзятое признание неизбежных предубеждений позволяет на ежедневных брифингах дополнять и уравновешивать восприятие сотрудников внешними участниками, которые, как известно, имеют противоположный опыт и предубеждения.
Устранение непродуктивного трения
Граница между наводящим на размышления опытом и неприятным конфликтом легко стирается. Для организации, которая так ценится за счет использования точек зрения из разнообразной человеческой сети, управление возникающими непродуктивными трениями имеет решающее значение. Межличностный конфликт, стыд, осуждение и неуважение могут быстро изменить окружающую среду, в результате чего члены рабочей группы менее склонны предлагать или оспаривать расходящиеся идеи или точки зрения.
Вне брифингов, при выборе сигналов, группа склонна создавать пространство для большего количества возможных точек зрения на брифинге, включая конкретный сигнал, если член решительно настроен по этому поводу, вместо того, чтобы обсуждать его включение. Результаты в виде комментариев, подключений и клиентских тегов для этого сигнала могут быть оценены позже и будут использоваться для выбора сигнала для следующего брифинга. Это укрепляет доверие в рабочей группе, потому что все участники соглашаются с объективной оценкой выбранных сигналов в отражении после действия.Объективные данные, общая методология и язык того, как они классифицируют и оценивают сигналы, встроенные в систему активного обучения S&H, помогают рассеять потенциально непродуктивные трения, которые могут возникнуть из-за разногласий относительно того, достаточно ли интересен конкретный сигнал для брифинга.
Чтобы помочь смягчить и развеять любые потенциально непродуктивные взаимодействия, которые могут истощить энергию и доверие, рабочая группа по брифингу поощряет лексику, которая является более конструктивной, чем конфронтационной, с формулировками, взятыми из проверенных ненасильственных коммуникационных стратегий, разработанных, чтобы помочь всем сторонам и точкам зрения быть услышанными.Например:
- «Я понимаю, откуда вы, но думаю, что это тоже интересно подумать. . . »
- «Да, в этом есть смысл. Я хочу развить эту мысль. . . »
- «Чтобы обеспечить обратную перспективу. . . »
В ежедневном брифинге непродуктивные трения — такие как стыд или осуждение — могут заставить участников почувствовать себя небезопасными или нежеланными, и они будут менее склонны к открытому участию. Конструктивная лексика стала еще более важной на брифингах.Например, при обсуждении сигнала о приложении, чтобы помочь профессионалам решить, следует ли им выступать на собрании, двое участников категорически не согласились с тем, является ли это положительным индикатором тенденции к «влиятельным женщинам». Третий участник сломал динамику поляризации, расширив дискуссию, сказав: «Я понимаю, откуда вы оба, хотя я хотел бы также рассмотреть это через призму культуры. . . » Это открыло дискуссию, чтобы участники могли искать дополнительные связи и интерпретации, а не застревать в противостоянии.
Рабочая группа также пытается уменьшить непродуктивные трения, поощряя использование невербальных сигналов, указывающих на одобрение или несогласие, и разрешая тихие беседы один на один, чтобы участники могли проявлять реакцию в данный момент, не прерывая группу в целом.
Не случайно ежедневный брифинг назначен на полдень, время, когда многие чувствуют, что их энергия иссякает. Многие участники находят мероприятие настолько заставляющим задуматься, что оно дает им заряд энергии, который поможет им провести остаток дня.Примечательно, что фасилитаторы рабочих групп работают над тем, чтобы создать «комфортный уровень дискомфорта», как выразилась директор человеческих сетей Аннали Киллиан, полагая, что борьба с новыми идеями требует определенного уровня дискомфорта.
Размышляйте больше, чтобы учиться быстрее
В промежутках между планированием и проведением ежедневных брифингов члены рабочей группы находят время, чтобы поразмышлять над брифингами, а также над тем, насколько эффективно консультант извлекает информацию из брифингов. Они также размышляют над любым новым представлением о том, где сигналы соответствуют культурной таксономии консалтинговой компании.Каждую неделю они рассматривают еженедельные изменения на брифингах и задают ключевые вопросы: Получили ли мы максимальную отдачу от нашего обсуждения какой-либо новой тенденции или ранее выявленной тенденции? Изменилась ли значимая ключевая тенденция?
Периодические брифинги, которые транслируются в прямом эфире, записываются и публикуются в социальных сетях, дают рабочей группе гораздо больше информации, которую нужно обдумать и использовать для настройки и повторной калибровки. Записи были особенно полезны для докладчиков, чтобы воспроизвести и проанализировать свою собственную работу, сравнить с метриками отслеживания и самокорректировать, чтобы улучшить скорость, четкость и настройку диалога для каждого сигнала, а также то, как они управляют и модерируют разговоры.Они также получают дополнительное представление об уровне вовлеченности и участия в сигналах, которые могут быть не очевидны во время проведения брифинга. Рабочая группа принимает в качестве входных данных обычные показатели вовлеченности и активности онлайн-аудитории и экспериментирует с тем, чтобы заставить онлайн-аудиторию предоставлять входные данные и проверять или оценивать сигналы, обсуждаемые во время брифингов в режиме реального времени.
На ежедневных брифингах цель состоит в том, чтобы охватить как можно больше территории, одновременно раскрывая как можно больше неожиданных и необычных идей.Учитывая эту проблему, рабочая группа разработала две метрики для отслеживания в качестве основных индикаторов количества и разнообразия связей, установленных в ходе брифинга, что является ключевым индикатором эффективности брифинга. Группа постоянно оценивает баланс между этими двумя показателями:
Комментариев на сигнал. Рабочая группа использует количество комментариев, сделанных при обсуждении сигнала, чтобы оценить, заходят ли участники слишком глубоко, что мешает брифингу охватить другие ценные сигналы или, наоборот, не проникнуть под поверхность, что препятствует выявлению закономерностей или раскрытию новых идеи.Количество комментариев также указывает на то, посчитали ли участники сигнал провокационным, что связано с тем, насколько глубина и нюансы нужно было распаковать, и какую потенциальную ценность этот сигнал может предложить клиентам. Клиенты, скорее всего, будут меньше ценить сигналы или идеи, которые являются прямыми или поверхностными.
Время, затраченное на сигнал. Среднее время, которое участники тратят на обсуждение любого сигнала, — еще один показатель, который помогает рабочей группе сбалансировать широту и глубину брифингов.
В результате размышлений над показателями и обсуждения конкретных брифингов рабочая группа разработала методы, призванные сделать брифинги более интересными и продуктивными. Например, в ходе брифинга каждый обсуждаемый культурный сигнал соединяется с другим в «сборки», которые продвигают разговор вперед. Группа обнаружила, что два-три сигнала на каждую сборку помогают поддерживать оптимальную динамику.
Помимо обсуждения индивидуальных сигналов, рабочая группа тратит время на установление связей между сигналами и между ними для определения возникающих культурных пересечений.Распознавание образов включает использование пяти метакатегорий: технологии и наука, человечество, идеология, эстетика и медиа, а также отслеживание того, как тенденции развиваются с течением времени. Участники, желающие работать с выводами, вытекающими из брифинга, записывают их на стикерах, а затем группируют их рядом с другими сигналами, с которыми они могут идентифицировать связь. Если образец становится более сильным, а его кластеры становятся достаточно большими, это определяется как «элемент культуры» (см. Рисунок 3).
Все эти прогнозы дополняются и усиливаются набором постоянно дорабатываемых алгоритмов, которые вместе показывают влияние любого заданного тренда путем измерения взаимодействия трех факторов: энергии, прогнозирования, и достижения .Энергия — это мера того, насколько быстро конкретный сигнал проходит через культуру. Прогнозирование — это оценка того, как долго тренд, в котором воплощен этот сигнал, может оставаться актуальным и / или развиваться с течением времени. Охват — это мера того, на скольких людей в настоящее время влияет тенденция или, по прогнозам, она повлияет со временем.
Комбинация этих трех показателей определяет и формирует место сигнала в таксономии тренда s&h и помогает определить связанный тренд как микро-, макро- или мега-тренд.Собственная методология оценки, разработанная консалтинговой компанией, определяет, движется ли тренд от микро к макро или от макро к мега, где охват, устойчивость и энергия определяют классификацию. Одним из примеров является рост популярности штанов для йоги как основной моды. То, что появилось сначала, помимо йоги, как легкая одежда для активных мам, в конечном итоге появилось в виде товаров от дизайнеров роскошной одежды; в одном брифинге s&h точно предсказала, что эта популярность на многих уровнях приведет к снижению покупок джинсовых джинсов в следующем году.
Процесс измерения также дает возможность выделить и сравнить измерения каждого тренда, например, чтобы определить, находится ли один на грани объединения с другим. Микротренды обычно обладают большей энергией, а макротренды более предсказуемы и достижимы.
Интервью «Дрэг-рейсинг РуПола»: Хани Дэвенпорт о том, как она превратилась из бездомной в работу на взлетно-посадочной полосе
Кристофер Рудольф
В эпизоде RuPaul’s Drag Race на этой неделе королевы привели нас в церковь, когда их разделили на команды телеевангелистов и попросили поклониться у алтаря поп-диве.Одна команда выбрала Бритни Спирс, а другая — Мэрайю Кэри. Но после доказательства того, что они не были частью ламбилий, РуПол заставил всех шести королев из команды Мэрайи синхронизировать губы друг с другом, в конечном итоге отправив Хани Дэвенпорт домой.
После ее исключения MTV News поговорила с Хани о ее времени в шоу, о том, на что была похожа эта историческая синхронизация губ, и как она превратилась из бездомной в работу на взлетно-посадочной полосе Drag Race .
Привет, дорогая! Я видел, как вчера вечером у вас была вечеринка в Нью-Йорке с Джинджер Миндж и Дасти Рэй Боттомс.Каково было устроить вечеринку, зная, что тебя отправят домой?
Это было действительно здорово! Меня окружало много родственников и друзей. Джинджер Миндж была там, и ее дважды исключали, так что она знает, каково это. Я боялся смотреть эту серию с тех пор, как вернулся со съемок сезона. [В то время] я чувствовал себя смущенным и не очень гордился собой, но потом, просмотрев его вчера вечером, я подумал: «О, на самом деле я не расстроен из-за этого». Я отдал все, и я сделал все, что мог.Я показал всему миру свое сердце. Этого было недостаточно, чтобы продвинуться дальше в сезоне, но на самом деле это был действительно хороший опыт.
На Untucked , вы рухнули на полу за кулисами после того, как РуПол устранил вас. Вы были очень расстроены. О чем вы думали в тот момент?
Валентина — не единственная, кто получает теленовеллу! [Смеется] Я был очень подавлен, потому что чувствовал, что все мои большие мечты о моей карьере заканчиваются прямо тогда, только чтобы понять, что это было только начало.
Во время испытания, когда вас попросили выбрать примадонну для поклонения, вы когда-нибудь думали выбрать другую примадонну, поскольку никто из вашей команды не был большим поклонником Мэрайи?
Мои товарищи по команде, казалось, были полностью согласны с Мэрайей, и я продолжал говорить: «Кто такая суперфанат Мэрайи Кэри?» [Я] думал, что раз уж мы выбрали ее, кто-то был, но это оказалось просто плохим решением.
РуПол довольно резко критиковал.Каково было находиться на сцене и слышать, как она говорит вашей команде, что вы «воняет»?
В тот момент я почувствовал себя таким неудачником, чего я просто не чувствую сегодня. Честно говоря, я так горжусь тем, чего мне удалось добиться. Я ехал на RuPaul’s Drag Race , я отлично выглядел, я хорошо себя показал. Может быть, я мог бы говорить больше, но я сделал все, что мог. Даже в этой синхронизации губ я дал 110%. Повествование не разыгралось там, где я выиграл, но я чувствую себя победителем.
Это еще начало сезона, поэтому ферзей много. Вы чувствовали, что вам не хватает экранного времени?
Еще до того, как я уехал на Drag Race, я имел дело с множеством. Один из моих наставников впал в кому, я был бездомным, и мне пришлось врезаться в кушетки друзей. Потом мне позвонили из Drag Race , и ты не говоришь маме «нет», когда она звонит. Мне просто не было места, чтобы соревноваться на моем высшем уровне. Так что я действительно в восторге от своего эфирного времени и от того, как меня показали.Когда я был там, мне пришлось многое пережить, и я просто пытался пережить это.
RuPaul заставил всю вашу команду синхронизировать губы друг с другом. Каково это было противостоять пяти другим ферзям одновременно? Вы спрыгнули со сцены!
Это был настоящий момент! Я бросился смертельно в халате, я не просто прыгнул на пол! Весь опыт был размытым. Я думал, что мне будет так больно, когда я посмотрю серию, но я просто очень горжусь всем, что я там сделал, и очень благодарен за возможность.
Наконец, можно поговорить о вашей новой песне «The Hive?» Это буп!
«The Hive» — это часть моего нового визуального альбома, в котором я буду носить то, что я буду носить на подиуме каждую неделю. Это не просто жестокая музыка, это социальный комментарий о том, каково быть цветным пидором в современной Америке. Я занимаюсь музыкой столько, сколько занимаюсь перетаскиванием, то есть уже 10 лет. Теперь, после Drag Race , я готов просто продолжать и продолжать творить.
RuPaul’s Drag Race выходит в эфир по четвергам в 9 / 8c на канале Vh2.
Последний вирусный тренд TikTok — джингл из российских хлопьев с 2010 года
Последняя вирусная тенденция TikTok уходит корнями в маловероятный источник: российскую рекламу хлопьев 10-летней давности.
На видео изображена анимированная компьютером лама, танцующая под завораживающий высокий вокал в пустыне, купаясь в апокалиптическом красном свете.
Детская песня — это ремикс на мелодию из рекламы русских хлопьев Miel Pops, или Honey Pops, 2010 года.
Его текст примерно переводится так: «Миэль Попс, гудение, гудение, гудение, жужжание. Миэль Попс, такая вкусная, ом ном ном. Медовые шарики на завтрак. Миэль Попс, капелька меда, капля капля, капля.
Возрождение джингла произошло, когда российская певица Черная Царевна загрузила видео, на котором она исполняет акустический кавер на эту песню.Другой пользователь ускорил звук и использовал его для озвучивания записи экрана, когда она играет в видеоигру.
Оттуда пользователь TikTok по имени awa_de_horchata_uwu взял ускоренный звук и использовал его в качестве саундтрека к танцующей ламе, запустив миллион мемов.
@awa_de_horchata_uwuMi😳pan🧟♀️su😎su🥳sum😡su👺su☠️su🤒mi😈pan💩yakakus🤖ñam👄ñam🙇🏼ñam
♬ ЭТА ПЕСНЯ НЕ О ХЛЕБЕ Stop mipansusus — itzmilpops
Многие испаноязычные пользователи слышали слова «mi pan» или «мой хлеб» вместо имени Miel Pops и ошибочно полагали, что этот джингл провозглашает любовь к хлебу.В результате во многих видео с использованием звука люди танцуют с буханками хлеба.
Рожденная из недр «alt TikTok», термин, относящийся к самым странным, жутким и сюрреалистичным уголкам видео-приложения, танцующая лама с тех пор вошла в массовое сознание поколения Z, набрав более 85 миллионов просмотров.
@jashlemNuestro pan 🥖 @ kimberly.loaiza
♬ ЭТА ПЕСНЯ НЕ О ХЛЕБЕ Stop mipansusus — itzmilpops
В то время, когда мало что в этом мире имеет смысл, неудивительно, что миллионы людей находят удовольствие в бессмысленном видео с жуткой антропоморфной ламой.
Это не первый случай, когда Россия становится маловероятной отправной точкой для популярных мемов. Этой весной группа танцоров из Ганы стала всемирно известной мемой после того, как видео их танца было смешано с русской электронной песней.
Понимаю, что происходит, но эта песня вроде пощечина 💀💀💀 pic.twitter.com/w1hSUHwbcs
— Джимми из LinkedIn (@LUSHHFOREVER) 1 августа 2020 г.
Распространение РНК-содержащих вирусов пчел у медоносной пчелы (Apis mellifera) в нескольких регионах России
Roulston, T.H. и Guodell, K, Роль ресурсов и рост в регулировании популяций диких пчел, Annu. Преподобный Энтомол. , 2011, т. 56. С. 293–312. doi 10.1146 / annurev-ento-120709–144802
CAS Статья PubMed Google ученый
Удина И.Г., Гришечкин А.Е., Кунижева С.С., Калашников А.Е., Учаева В.С., Злобин В.И., Кривцов Н.И., RT-PCR обнаружение вируса деформации крыла (DWV) у медоносной пчелы Apis mellifera L.в Московской области, Вопр. Virusol. , 2010, т. 55, нет. 5. С. 37–39. http://www.ncbi.nlm.nih.gov/pubmed/21260995.
CAS PubMed Google ученый
Удина И.Г., Гришечкин А.Е., Калашников А.Е., Злобин В.И., Кривцов Н.И. Идентификация вируса деформации крыла (DWV) у медоносной пчелы, Вестн. Росс. Акад. С-кх. НАУК .2010. 1. С. 69–71.
Google ученый
Ломакина Н.Ф. , Батуев Ю.М. Новый генотип вируса крестцово-подвздошных пчел Apis mellifera // Мол. Genet., Microbiol. Virol. , 2012, т. 27, нет. 3. С. 127–134. doi 10.3103 / S08
812030068
Статья Google ученый
Калашников А.Е., Масленников И.В., Колбина Л.М., Удина И.Г. Генетическая дифференциация популяций медоносной пчелы ( Apis mellifera L.) и распространение РНК-содержащих вирусов на фоне эпизоотий. Варроа деструктора на территории Удмуртии, С-х.Биол. .2013. 4. С. 88–92. doi 10.15389 / agrobiology. 2013.4.88eng. http://www.agrobiology.ru/4-2013kalashnikov-eng.html.10.15389/agrobiology.2013.4.88eng.
Google ученый
Дитеманн, В., Нацци, Ф., Мартин, С.Дж., Андерсон, Д.Л., Лок, Б., Делаплан, К.С. и др., Стандартные методы исследования Варроа, J. Apic. Res. , 2013, т. 52, нет. 1. С. 1–54. http://dx.doi.org/. doi 10.3896 / IBRA.1.52.1.09
Google ученый
Bereneyi, O., Bakonyi, T., Derakhshifar, I., Koglberger, H., Topolska, G., Ritter, W., et al., Филогенетический анализ генотипов вируса деформации крыла из различных географических регионов указывает на недавнее глобальное распространение вируса, заявл. Environ. Microbiol. , 2007, т. 73, нет. 11. С. 3605–3611. doi 10.1128 / AEM.00696-07
Артикул Google ученый
Берени О., Баконьи Н., Дерахшифар И., Коглбергер Х., и Новотны, Н., Встречаемость шести вирусов медоносных пчел на больных австрийских пасеках, Appl. Environ. Microbiol. , 2006, т. 72, нет. 4. С. 2414–2420. doi 10.1128 / AEM.72.4.2414-2420.2006
CAS Статья PubMed PubMed Central Google ученый
Де Миранда, Дж. Р., Кордони, Дж., И Бадж, Г., Вирус острого паралича пчелы-вирус пчелы Кашмира-вирус израильского острого паралича, J. Invertebr. Патол., 2010, т. 103, нет. 1. С. 30–47. doi 10.1016 / j.jip.2009.06.014
Статья Google ученый
Чен Ю., Эванс Дж. И Фельдлаунтер К. Горизонтальная и вертикальная передача вирусов у медоносной пчелы Apis mellifera, J. Invertebr. Патол. , 2006, т. 92, нет. 3. С. 152–154. doi 10.1016 / j.jip.2006.03.010
Статья PubMed Google ученый
Пиколл, Р. и Смаус, П.Е., GenAlEx 6.5: Генетический анализ в Excel. Программное обеспечение популяционной генетики для обучения и исследований — обновление, Bioinformatics , 2012, vol. 28, вып. 19. С. 2537–2539. DOI 10.1093 / биоинформатика / bts460
CAS Статья PubMed PubMed Central Google ученый
Антунес, К., Анидо, М., Гарридо-Байлон, Э., Ботиас, К., Зунино, П., Мартинес-Сальвадор, А. и др., Низкая распространенность вирусов медоносных пчел в Испания в 2006 и 2007 годах, Res.Вет. Sci. , 2012, т. 93, нет. 3. С. 1441–1445. doi 10.1016 / j.rvsc.2012.03.006
CAS Статья PubMed Google ученый
Антунес, К., Д’Алессандро, Б., Корбелла, Э. и Зунино, П., Обнаружение вируса хронического паралича пчел у уругвайской медоносной пчелы, J. Invertebr. Патол. , 2005, т. 90. С. 69–72. doi 10.1016 / J.JIP.2005.07.001
Статья PubMed Google ученый
Чен, М., Цуй, Л., Остиги, Н., и Кокс-Фостер, Д., Сложные пути передачи и взаимодействия между пикорноподобными вирусами (вирус пчелы Кашмира.