Ю-Мед, центр слуха в Санкт-Петербурге на метро Владимирская — отзывы, адрес, телефон, фото — Фламп
Уважаемая Лидия, мы внимательно изучили Ваши замечания, и, к сожалению, есть детали по поводу которых хотелось бы прояснить лично с Вами. В нашей фирме действительно работает опытный специалист Вышинская Светлана Васильевна, в слухопротезировании с 1982 года, имеет много положительных отзывов и благодарностей от наших клиентов. Также хочу отметить…
Показать целиком
Уважаемая Лидия, мы внимательно изучили Ваши замечания, и, к сожалению, есть детали по поводу которых хотелось бы прояснить лично с Вами. В нашей фирме действительно работает опытный специалист Вышинская Светлана Васильевна, в слухопротезировании с 1982 года, имеет много положительных отзывов и благодарностей от наших клиентов. Также хочу отметить тот факт, что с 2006 года она НИКОГДА не совмещала данную работу с практикой лор-врача в лечебных учреждениях, таким образом она не могла направить Вас из поликлиники к нам в Центр слуха. Мы пытались найти Вашу карту в нашей регистратуре, но за последние 6 месяцев у Вышинской Светланы Васильевны не было ни одного первичного клиента по имени Лидия. Если Вы обратитесь к нам напрямую, мы сможем поднять данные, Вашу карту, соблюдая полную конфиденциальность, и восстановить ситуацию, а также понять какие конкретные варианты Вам предлагал специалист. По нашему внутреннему стандарту предлагается на выбор 3 варианта и обычно разной ценовой категории, а не один слуховой аппарат. К нам приходят клиенты из разных источников, в том числе по рекомендации врачей разных медицинских центров, для нас это большая честь и доверие, которое мы стараемся оправдывать. Именно для того, чтобы человек был удовлетворён, у нас у единственных в городе уже 7 лет есть ГАРАНТИЯ на возврат или обмен слухового аппарата в течение 3 месяцев, таким образом, мы можем быть уверены, что наши клиенты, покупая слуховые аппараты СЛЫШАТ.
Центр слуха Ю-мед, слуховые аппараты, Россия, Санкт-Петербург, Большая Московская ул., 16 — Яндекс.Карты
Моей дочке Софии 10 лет, у нас 3 степень сенсоневральной тугоухости. Аппаратики мы носим с 3-х лет и уже успели перепробовать несколько фирм и моделей. Нам очень повезло найти своего специалиста — Анна Александровну Ефимову, в центре слуха Ю-МЕД. Знаете, это удивительное место, где концентрация тепла и душевных людей, просто зашкаливает! В центе слуха Ю-мед, о слухе моего ребёнка правда заботятся, а еще о том, чтобы напоить чаем, участливо напомнить о предстоящем приеме, встретить с доброжелательной улыбкой! Побольше бы таких мест, где специалист в халате, скорее друг, чем сторонний наблюдатель. Анна Александровна сумела подобрать к моему ребёнку ключик и Софийка бежит на каждый прием, как на праздник! И вот в феврале, мы стали обладателями Phonak Marvel Audeo 90-R и тут все, что мы знали ДО перевернулось. Дочка не могла сдержать эмоций. Первое, что она сказала одев новые Марвеллы, «мама я слышу как шуршат бахилы». Идя по улице она восклицала сколько вокруг звуков и почему она их не слышала ДО. Так как звук образуется ближе к уху, практически в нем, широта звучания просто поражает. Что я могу сказать, как мама, про это новейшее слово техники. Про этот мини компьютер на ушах моего ребёнка. Дома стало тише, теперь мультики Софийка смотрит напрямую подключаясь через блютус к телефону или планшету.
FLEX:TRIAL от Unitron – список клиник, участвующих в акции
1. Центр ЛОР-помощи и Слухопротезирования
Самара, ул. Галактионовская 106, А (2-й этаж)
Tel: +7 927 732-0762
www.lorinfo.ru
2. Городской центр слуха
Киров, ул. Ленина, дом № 89/1, «Кабинет слухопротезирования»
Tel: +7 922 975-14-26
хорошийслух.рф/
3. Городской Центр Коррекции Слуха «Абсолютный Слух»
Барнаул, ул. Союза Республик, 36 Б, оф. 24
Тел.: +7 952 138 04 88
sluh32.ru
4. Центр слуха Ю-МЕД
Санкт-Петербург, ул. Большая Московская, дом 16, литера А, пом.5Н
Тел.: 8 812 575-76-26, 812 570-05-69
www.you-med.ru
5. Центр «Ото Плюс»
Иркутск, мкрн Юбилейный, д. 100, офис 806
Тел.: 8 (3952)951-079, (3952)951-019
6. Центр коррекции слуха Лори (ООО «Стоматех-Л»)
в Екатеринбурге:
Ул. Тимирязева 13
Тел.: 8-343-375-85-76
Ул. Белореченская 29
Тел.: 8-343-31786-66, 8-900-199-13-99
Ул. 8Марта 142
Тел.: 8-343-263-60-63, 8-900-204-15-40
Ул. Машинострителей 49
Тел.: 8-343-338-00-17, 8-922-294-22-62
в Нижнем Тагиле:
Ул.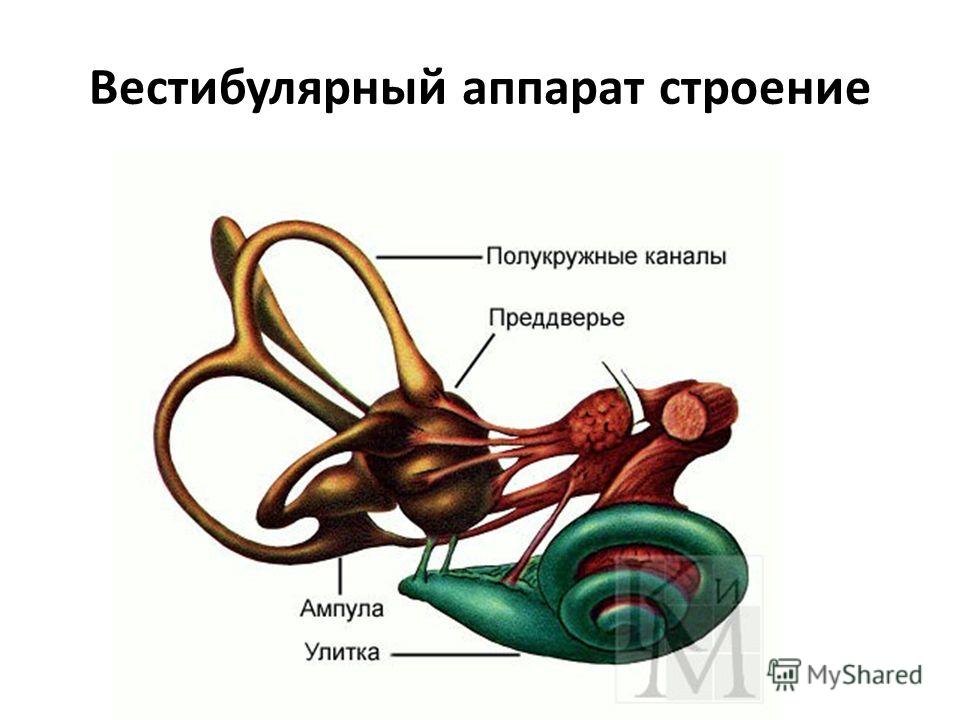 К. Маркса 64/Окт револ. 31
К. Маркса 64/Окт револ. 31
Тел.: 8-3435-25-7517, 8-922-100-99-85
в Каменск-Уральском:
Ул. Победы 39
Тел.: 8-3439-545-000, 8—952-14-88-750
в Тюмени:
ул. 8 Марта 1(вход с ул. Советская)
Тел.: 8-3452-230-130, 8-922-079-40-77
в Челябинске:
Ул. Красная, 63-оф. 11
Тел.: 8-351-266-19-75, 247-67-17(8-922-710-67-17)
Ул. Арзамасская 5-каб.2
Тел.: 8-351-225—27-32, 8-952-527-60-13
в Кургане
ул. Красина 53 оф 6/1
Тел.: 8-3522-41-62-57, 62-66-66
ул. Ленина 44
Тел.: 8-3522-233-060, 8-912-835-0503
в Миассе:
пр. Автозаводцев , 15
Тел.: 8-3513-55-01-23, 8-932-309-39-32
в Златоусте:
ул. Карла Маркса 19
Тел.: 8-3513-690-217, 8-932-015-16-16
в Новоуральске:
ул. Юбилейная, д.3
Тел.: 8-34370-5-85-37, 8-932 112 43 77
7. Центр коррекции слуха и речи «Мелфон»
в Москве:
ул. Профсоюзная, д.78, стр.1,
Тел.: +7 (495) 258-35-13
www.melfon.ru
в Санкт-Петербурге:
ул. Московский проспект, дом 117А,
www.melfon.ru/spb
в Самаре:
ул. Киевская, д. 13 (вход во двор напротив храма)
Тел.: 8-927-725-37-02, 8 (8462) 47-77-75
www.melfon.ru/samara
в Воронеже:
проспект Патриотов, д. 53-A
Тел.: 8 (4732) 31-86-55, 91-58-15
http://melfon-vrn.ru/
отзывы, адрес, время работы, расположение на карте, посещаемость
Медицинская клиника Медицинский центр слуха Ю-МЕД, Большая Московская ул., 16, Санкт-Петербург, Ленинградская обл., 191002: 29 отзывов пользователей и сотрудников, подробная информация о адресе, времени работы, расположении на карте, посещаемости, фотографии, меню, номер телефона и огромное количество другой подробной и полезной информации , 16, Санкт-Петербург, Ленинградская обл., 191002
, 16, Санкт-Петербург, Ленинградская обл., 191002Сайт: you-med.ru
Номер телефона: 8 (812) 570-05-69
Расположение на карте
Время работы
| Понедельник | 09:00–19:00 |
| Вторник | 09:00–19:00 |
| Среда | 09:00–19:00 |
| Четверг | 09:00–19:00 |
| Пятница | 09:00–19:00 |
| Суббота | 10:00–18:00 |
| Воскресенье | Закрыто |
Отзывы
Ска СероваОчень хороший медицинский центр! Врачи профессионалы! Администраторы вежливые, предупредительные. Чистота в кабинетах безупречная. Соблюдаются все нормы карантина. Я довольна всем!
9 месяцев назад
Маришка Гусева
Расскажу свою историю. Обратилась в этот центр по рекомендации после операции. Левое ухо после тимпанопластики не слышит, как результат трудности с общением и работой. В конце февраля 2019 заказала внутриушной аппарат за 30 000 р. марки Фонак. Получила его 02.03.2019, обслуживала добросовестно, сурдолога посещала ежемесячно и показывала и ухо и аппарат. Регулярно меняла фильтр, но 05.03.2020 т.е. ровно через год аппарат сломался. Ремонт оказался вовсе не гарантийный, якобы ресивер забит серой, хотя врача посетила накануне 09.02.2020. Стоимость ремонта составила 9 000 р., после моего возмущения была предложена скидка до почти 8 000 р. Если решите обращаться в этот центр подумайте, кофе и приветливый персонал вовсе не главное, надо на другие вещи обращать внимание, но это мой первый опыт. с 03.03.2020 я пенсионерка, для меня и аппарат и его ремонт большие деньги. Я очень разочарована.
Обратилась в этот центр по рекомендации после операции. Левое ухо после тимпанопластики не слышит, как результат трудности с общением и работой. В конце февраля 2019 заказала внутриушной аппарат за 30 000 р. марки Фонак. Получила его 02.03.2019, обслуживала добросовестно, сурдолога посещала ежемесячно и показывала и ухо и аппарат. Регулярно меняла фильтр, но 05.03.2020 т.е. ровно через год аппарат сломался. Ремонт оказался вовсе не гарантийный, якобы ресивер забит серой, хотя врача посетила накануне 09.02.2020. Стоимость ремонта составила 9 000 р., после моего возмущения была предложена скидка до почти 8 000 р. Если решите обращаться в этот центр подумайте, кофе и приветливый персонал вовсе не главное, надо на другие вещи обращать внимание, но это мой первый опыт. с 03.03.2020 я пенсионерка, для меня и аппарат и его ремонт большие деньги. Я очень разочарована.
1 месяц назад
Александр БелецкийВсё супер! И персонал, и внимание к клиентам!
3 месяца назад
Александра Петрова
Персонал приветлив и добро желателен, на этом, пожалуй, все. Заказывала гидроплаги, сделали — в уши все равно попадала вода, переделали-с таким же результатом. Выброшенные на ветер три тысячи.
Выброшенные на ветер три тысячи.
9 месяцев назад
Наташа СаломатинаХорошие и внимательное обслуживание!
6 месяцев назад
Игорь МироновРекомендую, работают профи
6 месяцев назад
Юлька НеисправимаяЗамечательный медицинский центр! Маме подобрали слуховые аппараты, мне больше не нужно повышать голос, чтобы быть услышанной. Очень удобно, что можно взять аппараты на пробу на две недели и поменять, если не понравились в процессе эксплуатации. Со вкусом оформлен офис, персонал приветлив и доброжелателен. Оценка «пять» по всем пунктам.
11 месяцев назад
Юлия Пимишкова
Уютный центр слуха с отличным персоналом. Кившик Мария Юрьевна — отличный детский слухопротезист! Впечатления от центра только положительные! Ходим туда часто.
Кившик Мария Юрьевна — отличный детский слухопротезист! Впечатления от центра только положительные! Ходим туда часто.
2 месяца назад
Эдуард КочетковОтличное обслуживание.доброжелательная обстановка.
5 месяцев назад
Мария АгаповаХороший центр, приветливый персонал на ресепшене и настоящие профессионалы врачи. Помогли в решении моей проблемы.
9 месяцев назад
Анна ДегтяреваЗамечателный доктор Говорухина Ирина.
8 месяцев назад
Игорь МихайловРеспект этим людям
5 месяцев назад
Таисия КочетковаХорошие докторе и специальносты
5 месяцев назад
Дмитрий Васильев
Все на высшем уровне.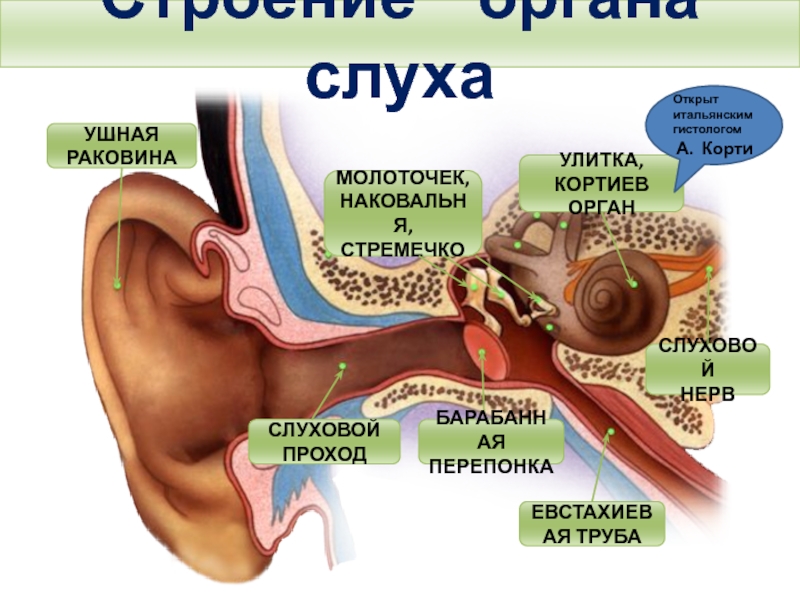
9 месяцев назад
Александр Виноградов8 месяцев назад
Мария Разумова11 месяцев назад
Александра Лесниченко10 месяцев назад
Иван Петров8 месяцев назад
Алексей Бороденков9 месяцев назад
Наталия Суренкова5 месяцев назад
Irina Fateycheva7 месяцев назад
Надежда Гончарова7 месяцев назад
Таалай Джумабаев7 месяцев назад
Алена Федотова1 месяц назад
Екатерина Салтыкова8 месяцев назад
Николай Дегтярёв8 месяцев назад
Даша Хрисаниди6 месяцев назад
Eight-Three Five-O11 месяцев назад
Котёнок Пушистый6 месяцев назад
Популярные места из категории Медицинская клиника
Расписание приема специалистов МЦ МастерСлух, Краснодар
Расписание приема
Прием в клиниках «МастерСлух» осуществляется по предварительной записи по телефонам
В расписании могут быть изменения, уточняйте у администратора клиники.
г.Краснодар, Красная, 154.
| Специалист | Понедельник | Вторник | Среда | Четверг | Пятница | Суббота | Воскресенье |
|---|---|---|---|---|---|---|---|
| Гауфман В.Е. | Прием ведется по предварительной записи | выходной | |||||
| Смирнова И.А. |
9:00-17:00 |
9:00-17:00 |
9:00-17:00 |
9:00-17:00 |
9:00-17:00 |
1 р. в месяц |
выходной |
| Смирнов П.В. |
9:00-17:00 |
9:00-17:00 |
9:00-17:00 |
9:00-17:00 |
9:00-17:00 |
1 р. в месяц |
выходной |
| Мержоев Т.М. |
10:30-18:00 |
10:30-18:00 |
10:30-18:00 |
10:30-18:00 |
10:30-18:00 |
1 р. в месяц |
выходной |
г. Краснодар, 1 мая, 186.
| Специалист | Понедельник | Вторник | Среда | Четверг | Пятница | Суббота | Воскресенье |
|---|---|---|---|---|---|---|---|
| Михайлова О.А. |
9:00-17:00 |
9:00-17:00 |
Нет приема |
9:00-17:00 |
9:00-17:00 |
2 р. |
выходной |
| Лазарева Ю.А. | Нет приема | 9:00-17:00 | Нет приема | 9:00-17:00 | Нет приема | 2 р. в месяц 9:00-17:00 |
выходной |
 в месяц
в месяцКраснодар, Железнодорожная, 2/1.
| Специалист | Понедельник | Вторник | Среда | Четверг | Пятница | Суббота | Воскресенье |
|---|---|---|---|---|---|---|---|
| Гауфман В.Е. | Прием ведется по предварительной записи | выходной | выходной | ||||
| Черняк Г.В. | 9:00-17:00 | 9:00-17:00 | 9:00-17:00 | 9:00-17:00 | Нет приема | выходной | выходной |
| Рылова Г.М. |
9:00-17:00 |
9:00-17:00 |
9:00-17:00 |
9:00-17:00 |
9:00-17:00 |
выходной |
выходной |
| Ютман К.Ю. | Нет приема | 9:00-17:00 | Нет приема | 9:00-17:00 | Нет приема | Нет приема | выходной |
г.Новороссийск, проспект Ленина, 23.
| Специалист | Понедельник | Вторник | Среда | Четверг | Пятница | Суббота | Воскресенье |
|---|---|---|---|---|---|---|---|
| Гауфман В.Е. |
Прием ведется по предварительной записи |
выходной |
выходной |
||||
Черняк Г. В. В. |
Прием ведется по предварительной записи | выходной | выходной | ||||
г.Армавир, ул.Мира, д.24, 4 этаж, офис 403
| Специалист | Понедельник | Вторник | Среда | Четверг | Пятница | Суббота | Воскресенье |
|---|---|---|---|---|---|---|---|
| Болотова Е.А. |
9:00-17:00 |
9:00-17:00 |
9:00-17:00 |
9:00-17:00 |
9:00-17:00 |
выходной |
выходной |
| Гауфман В.Е. | Прием ведется по предварительной записи | выходной | |||||
| Черняк Г.В. | Прием ведется по предварительной записи | выходной | |||||
Аудиометрия (измерение остроты слуха) | Медицинский центр «Президент-Мед»
Аудиометрия – современная методика диагностики, позволяющая оценить работоспособность слуховой системы человека и определить наличие различных нарушений органа слуха.
Аудиограмма не имеет противопоказаний, поэтому исследование может назначаться даже новорожденным детям. Диагностика помогает выявить патологии всех структурных отделов уха. Обязательно показана методика перед процедурой слухопротезирования.
Исследование остроты слуха производится посредством специального аппарата – аудиометра. С помощью такого медицинского оборудования врачи-оториноларингологи могут определить нижнюю и верхнюю границу слышимости пациента.
Возможности аудиометрии:
- Позволяет исследовать слуховую активность.
- Определяет восприятие речи.

- Обнаруживает нарушения слуха.

В обязательном порядке аудиограмма выполняется, если пациент жалуется на боль в ухе или на резкое снижение слуха. Показанием к проведению процедуры также является отит, состояние хронической или острой глухоты. Может назначаться больным для контроля результата пройденной терапии.
Виды аудиометрии
В зависимости от способа диагностики и применяемой во время исследования аппаратуры аудиометрия бывает речевой, тональной, а также компьютерной. Самым простым методом является речевой тип обследования, поскольку он проводится без использования специального оборудования.
Речевая аудиометрия производится во время первого осмотра пациента, а также для подтверждения врачебного диагноза или при подборе слухового аппарата. Для выявления остроты слуха используется разговорная речь и шепот. Методика довольно проста, но при этом она не дает точно установить степень понижения слуха.
Самым объективным вариантом диагностики слуха является компьютерная аудиометрия. Такое исследование не имеет противопоказаний, поэтому может назначаться маленьким детям, а также пациентам, которые перенесли травмы головы, инсульты и больным с нарушениями психики.
Тональная аудиометрия довольно информативна, она позволяет определить, какое ухо плохо слышит или вовсе не реагирует на звуки. Исследование помогает выявить порог чувствительности человеческого слуха к различным по частоте звуковым волнам. Результаты диагностики отображаются в аудиограмме, которая показывает, насколько звуковосприятие пациента отличается от нормы и на каких частотах происходят сбои.
Как проходит аудиометрия
Довольно часто пациентам для проверки слуха врач-оториноларинголог назначает тональную аудиометрию. Больному предлагают пройти в изолированную от внешних звуков кабину. Для исследования воздушной проводимости нужно надеть наушники, в которые будут подаваться звуки различной громкости и частоты. Диагностика каждого уха происходит отдельно, если человек что-то слышит, он должен нажать на кнопку, находящуюся перед ним. К недостаткам метода относят чувство дискомфорта, его пациенты могут испытывать на определенных звуковых частотах.
К недостаткам метода относят чувство дискомфорта, его пациенты могут испытывать на определенных звуковых частотах.
Во время речевой аудиометрии больному на расстоянии шести метров шепотом произносят определенные слова и словосочетания, он должен их повторить. При этом аудиограмма не чертиться, потому что точно определить разницу между чувствительностью к звукам правого и левого уха невозможно.
При компьютерном исследовании к голове больного прикрепляют электроды, по которым направляются щелчки и другие звуки различной интенсивности. Эти проводники соединены с компьютером. Во время диагностики доктор будет наблюдать за безусловными рефлексами пациента: расширением зрачков, морганием, пульсом и т. п. Все изменения, происходящие под воздействием акустической стимуляции в органах слуха, фиксируются на аппарате.
Для измерения остроты слуха обращайтесь в медицинские центры «Президент-Мед» в Москве (на Коломенской и м.ВДНХ) и в Видном
Автор: Лаврова Нина Авенировна
Заместитель генерального директора по медицинской части
Окончила Ярославский государственный медицинский институт по специальности «Лечебное дело»
Медицинский опыт работы — 25 лет
Записаться к врачу
ОТЗЫВЫ КЛИЕНТОВ
Ольга
Спасибо Юлие Владимировне! Врач очень опытная, внимательная, а самое главное ее очень любят дети! Приходилось уже несколько раз с ребёнком обращаться и всегда каждый приём на высоте. Как хорошо, когда есть такие хорошие врачи[…]Щетининой О.Н.
Большое спасибо всем работникам клиники за слаженную работу и хорошее настроение. Отдельное спасибо Ивановой Ю.А. за профессионализм, отзывчивость, индивидуальный подход.[…]«Проблемы возрастной тугоухости» | Городской центр медицинской профилактики
врач-сурдоакустик центра слуха Ю-МЕД Василий Николаевич Коробицин
По данным статистики, в России каждый шестой страдает от тугоухости, что составляет более 16% населения. Причем ослабление слуха может встречаться в любом возрасте, а не только у пожилых людей. Слуховая способность человека меняется в течение жизни. Максимальная острота слуха достигается в возрасте 25 лет, а затем медленно и незаметно начинает снижаться. С возрастом потеря слуха наблюдается чаще у мужчин, чем у женщин. В 60-70 лет двое из четверых мужчин имеют снижение слуха, а у женщин только у 1-2 из четверки.
Причем ослабление слуха может встречаться в любом возрасте, а не только у пожилых людей. Слуховая способность человека меняется в течение жизни. Максимальная острота слуха достигается в возрасте 25 лет, а затем медленно и незаметно начинает снижаться. С возрастом потеря слуха наблюдается чаще у мужчин, чем у женщин. В 60-70 лет двое из четверых мужчин имеют снижение слуха, а у женщин только у 1-2 из четверки.
Тугоухость – это снижение слуха, затрудняющее речевое общение. При этом только в 10% всех случаев потери слуха возможно лечение. Остальные 90%, благодаря развитию современных технологий, корректируются с помощью слуховых аппаратов.
Каковы причины, вызывающие потерю слуха? Главная — акустические нагрузки, вызванные шумом цивилизации. Отрицательную роль играют злоупотребление никотином, сосудистые изменения с нарушением кровообращения, последствия инфекции, особенности питания, наследственность, употребление некоторых лекарственных препаратов.
Частое переспрашивание в разговоре, потребность в увеличении громкости радио или телевизора, трудности в определении источника звука, затруднение при разговоре в компании, когда говорят сразу несколько человек, ухудшение разборчивости речи в шумной обстановке – все это признаки снижения слуха. Но мы часто не обращаем на них внимание. А ведь чем раньше мы выявим снижения слуха, тем успешнее сможем компенсировать эту потерю.
Запомните, что начиная с 50 лет проверка слуха должна стать частью вашего регулярного профилактического осмотра.
Своевременное аудиологическое обследование позволит обнаружить даже минимальное снижение слуха. У пациентов с выявленной тугоухостью различной степени контроль слуха должен осуществляться раз в год.
Когда снижение слуха становится очевидным, необходима консультация лор – врача. Этот специалист исключит случаи, требующие хирургического вмешательства, классифицирует нарушение слуха как подлежащее или не подлежащее коррекции, определит степень потери слуха и показания к слухопротезированию, направит к врачу — сурдологу.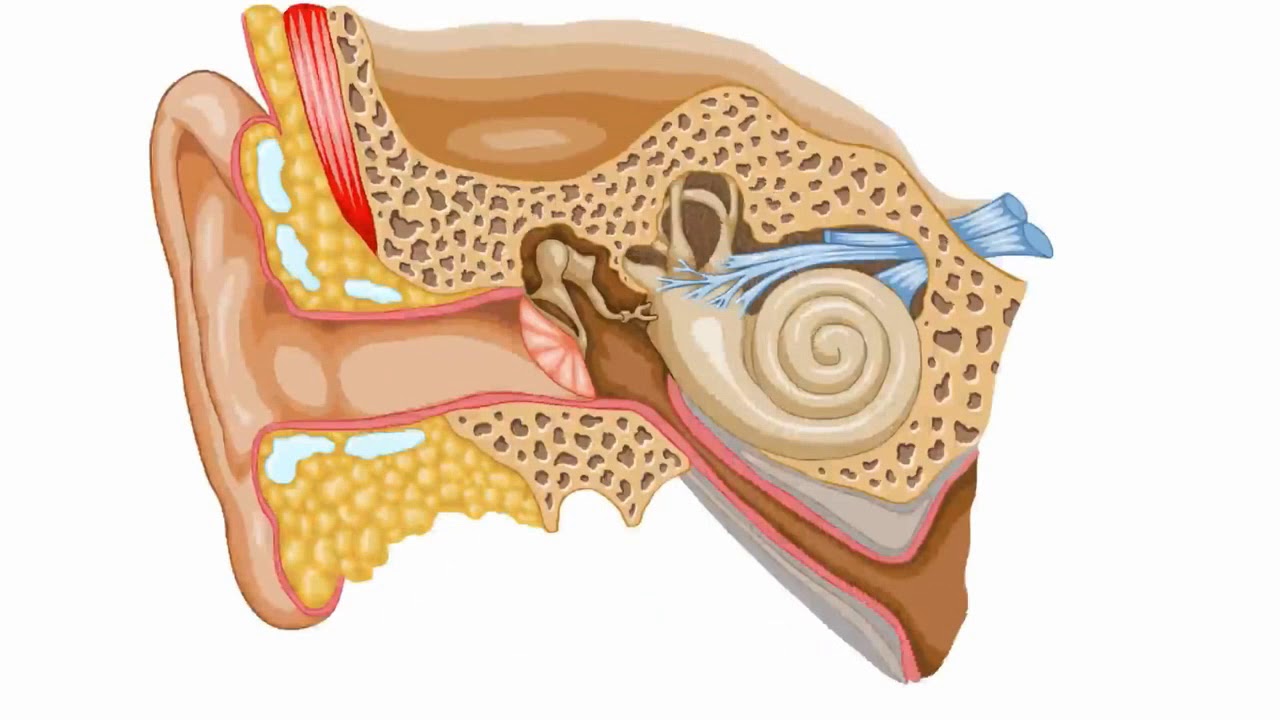 В свою очередь, сурдолог выяснит жалобы и нужды пациента, сделает слепок уха и проведет необходимые исследования – отоскопию, тональную и речевую аудиометрию, Если сурдолог по результатам обследования поставит диагноз «тугоухость», то будет решаться вопрос о слухопротезировании. При этом человек сам решает, носить ему или не носить слуховой аппарат на основании своей слуховой потребности и слуховой ситуации. Но, делая выбор, человек со сниженным слухом должен помнить, что слуховой аппарат гораздо лучше, чем отторжение от социального окружения и изоляция. Не зря же говорят, что слепота отделяет человека от вещей, а снижение слуха отделяет его от людей. Человек, который не признает своей тугоухости и не пытается с ней бороться, обрекает себя на одиночество.
В свою очередь, сурдолог выяснит жалобы и нужды пациента, сделает слепок уха и проведет необходимые исследования – отоскопию, тональную и речевую аудиометрию, Если сурдолог по результатам обследования поставит диагноз «тугоухость», то будет решаться вопрос о слухопротезировании. При этом человек сам решает, носить ему или не носить слуховой аппарат на основании своей слуховой потребности и слуховой ситуации. Но, делая выбор, человек со сниженным слухом должен помнить, что слуховой аппарат гораздо лучше, чем отторжение от социального окружения и изоляция. Не зря же говорят, что слепота отделяет человека от вещей, а снижение слуха отделяет его от людей. Человек, который не признает своей тугоухости и не пытается с ней бороться, обрекает себя на одиночество.
Многие люди, нуждающиеся в слухопротезировании, не пользуются слуховыми аппаратами. Почему? У одних – это психологический барьер: «Я еще не старик, чтобы носить слуховой аппарат. Он бросается в глаза. Мне неловко его носить». У других — позднее выявление нарушения слуха, позднее применение слухового аппарата. У третьих — отрицательный опыт предыдущего использования слухового аппарата плохого качества, неправильно подобранного или настроенного. Вывод напрашивается сам собой – необходим профессиональный индивидуальный подбор слухового аппарата. Тем более, что сегодня для компенсации потери слуха предлагаются различные модели слуховых устройств. Большинство аппаратов носят за ухом. И хотя они довольно миниатюрны, некоторые стесняются их носить. В этом случае врач может подобрать внутриушной или даже внутриканальный аппарат, о котором будет знать только сам пациент. Для того, чтобы получить желаемый эффект, необходимо не только подобрать качественный аппарат, но и специально его настроить для конкретного пациента, так как наша слуховая способность так же индивидуальна, как и отпечатки пальцев. В противном случае, он будет действовать только как усилитель звуковых колебаний из внешней среды, а разрушение слуха продолжится.
Особо надо отметить, что и какие БАДы, лекарства и диеты не помогут вам вернуть полноценный слух. Не верьте рекламе. В случае тугоухости поможет только слуховой аппарат.
Медоносные пчелы избирательно избегают трудного выбора
Значимость
Здесь мы показываем, что медоносные пчелы ( Apis mellifera ) могут адаптивно изменять свое поведение в тесте выбора в ответ на трудность испытания. Пчелы предпочитают отказываться от сложных испытаний и тем самым повышают свой шанс на успех. Мы обсуждаем, включает ли этот выбор оценку степени неопределенности (рассматриваемой как определение базового метапознания) или эта задача может быть решена с помощью ассоциативных механизмов.Мы предлагаем гипотезу о том, как неопределенность может быть обработана в известных схемах мозга насекомых, чтобы сформулировать концепцию неопределенности как тему для нейробиологического анализа.
Abstract
На стратегии принятия решений человеком сильно влияет осознание уверенности или неопределенности (форма метапознания), что увеличивает шансы сделать правильный выбор. Люди ищут больше информации и откладывают выбор, когда понимают, что у них недостаточно информации для принятия точного решения, но осведомлены ли животные о неопределенности, в настоящее время очень спорный вопрос.Чтобы изучить этот вопрос, мы изучили, как медоносные пчелы ( Apis mellifera ) реагировали на задачу визуального различения, сложность которой варьировалась между испытаниями. Свободно летающие пчелы награждались за правильный выбор, наказывались за неправильный выбор или могли избежать выбора, покинув испытание (отказ от участия). Пчелы чаще отказывались от участия в сложных испытаниях, и отказ от них увеличил долю успешных испытаний. Пчелы также могут передать идею отказа от новой задачи. Наши данные показывают, что пчелы избирательно избегают сложных задач, для решения которых у них отсутствует информация.Это открытие было рассмотрено как доказательство того, что животные, не являющиеся людьми, могут оценить достоверность предсказанного результата, а производительность пчел была сопоставима с производительностью приматов в аналогичной парадигме.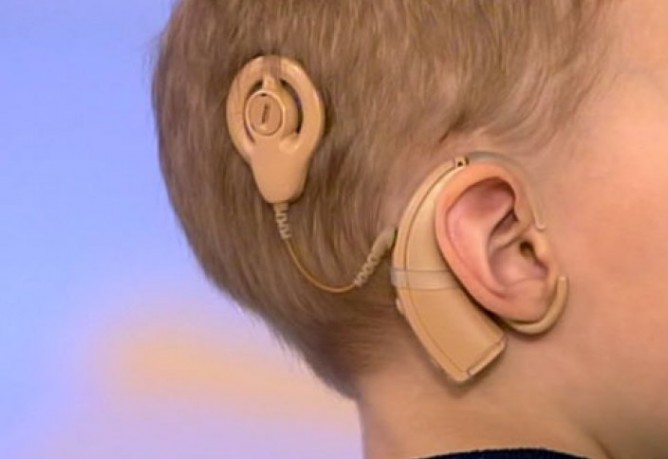 Мы обсуждаем, доказывают ли эти поведенческие результаты реакцию пчел на неопределенность или ассоциативные механизмы могут объяснить такие результаты. Чтобы лучше представить метапознание как проблему для нейробиологического исследования, мы предлагаем нейробиологическую гипотезу мониторинга неопределенности, основанную на известной схеме мозга медоносной пчелы.
Мы обсуждаем, доказывают ли эти поведенческие результаты реакцию пчел на неопределенность или ассоциативные механизмы могут объяснить такие результаты. Чтобы лучше представить метапознание как проблему для нейробиологического исследования, мы предлагаем нейробиологическую гипотезу мониторинга неопределенности, основанную на известной схеме мозга медоносной пчелы.
Часто правильный выбор можно только оценить, а не знать абсолютно. Чтобы помочь в этой оценке, люди могут отслеживать степень своей неопределенности и использовать эти знания для улучшения своих решений (1, 2). Способность контролировать собственные когнитивные процессы считается формой метапознания (1, 2). В случае сомнений люди часто откладывают выбор и ищут больше информации, чем рискуют последствиями неправильного выбора. Вопрос о том, существует ли возможность отслеживать неопределенность у нечеловеческих животных, в настоящее время вызывает большие споры.Smith et al. (3, 4) разработали парадигму отказа, чтобы проверить мониторинг неопределенности на животных, в котором животное должно решить задачу распознавания, которая различается по сложности. Животное награждается за правильный выбор и наказывается за неправильный выбор. Затем вводится третий вариант, при котором животное может «отказаться», ответив каким-либо другим способом, чтобы избежать задачи распознавания, тем самым обычно начиная новое испытание. Если животные отказываются больше от сложных, чем от легких задач, если отказ улучшает выполнение сложных задач, и если они могут применить стратегию отказа к новой задаче, то это было воспринято как доказательство того, что животные могут изменять свое решение. стратегия, основанная на степени их неопределенности.Этот результат был зарегистрирован для нечеловеческих приматов, дельфинов, собак и крыс (3⇓⇓⇓⇓⇓⇓⇓⇓ – 12). Однако некоторые решительно утверждают, что все сравнительные исследования с использованием парадигм отказа можно объяснить с помощью ассоциативных механизмов, которые не требуют суждений о неопределенности (3, 13–15).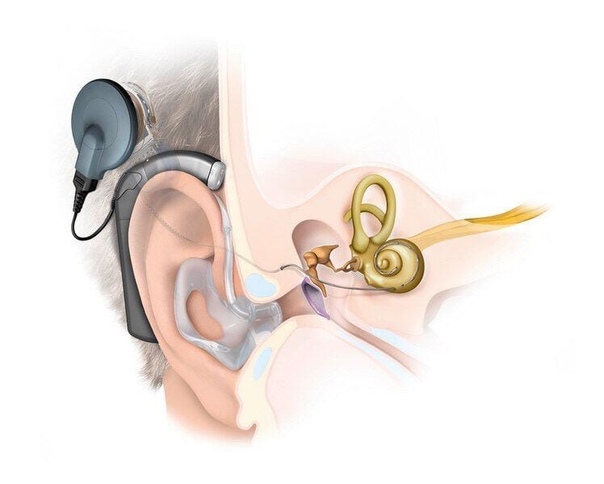
Поскольку были предложены два альтернативных механизма для объяснения одних и тех же поведенческих данных, Морган Канон (16) предупреждает, что при представлении двух альтернативных объяснений мы вынуждены выбирать более простой из двух.Однако прямое сравнение этих гипотез затруднено, поскольку не существует согласованной нейронной модели для обработки неопределенности. Поэтому сложно судить, какое объяснение действительно проще. Здесь мы вносим свой вклад в дискуссию, исследуя, как беспозвоночное, медоносная пчела, действует в рамках парадигмы отказа. Мы обнаружили, что пчелы соответствовали всем необходимым критериям, которые считаются показательными для мониторинга неопределенности. У пчел мозг намного меньше, чем у любого млекопитающего, и мы предполагаем, как это животное может обеспечить мониторинг неопределенности.
Результаты
Чтобы оценить, как медоносные пчелы реагировали на выбор, основанный на ограниченной информации, мы исследовали их производительность в задаче визуального различения, в которой сложность задачи варьировалась. Мы объединили задачу конфигурационного обучения, в которой пчелы должны были изучить стимулы, расположенные выше или ниже референта (17), с возможностью отказаться от испытаний (5), чтобы определить, изменили ли пчелы адаптивно поведение в ответ на сложность задачи.
Мы обучили свободно летающих пчел входить в двухкамерную испытательную установку (рис.1). Во время обучения стимулы помещались либо в первую камеру принятия решения, либо во вторую. Во время тестирования обе камеры содержали раздражители. Стимулы представляли собой две переменные цели, которые располагались либо над, либо под контрольной полосой (см. Ссылку 17) (рис. 1 B ). Одна группа пчел была обучена тому, что указанная выше цель содержала 2 М сахарозы (вознаграждение), а указанная ниже цель содержала 50 мМ хинина (наказание) (18). Для второй группы случайность была обратной. На этом этапе пчел тренировали с пятью разными мишенями, которые различались по форме, размеру и цвету. Мы изменили цели и вертикальное положение целей и контрольных полос псевдослучайно, чтобы исключить возможное использование ассоциативных сигналов. Другими словами, форма, цвет и размер мишеней менялись от испытания к испытанию, а расстояние до мишеней от дна камеры и относительно контрольных стержней варьировалось от испытания к испытанию. Этот формат гарантировал, что низкоуровневые сигналы, такие как совокупная область (область, охватываемая как контрольной полосой, так и целью) или конфигурационная идентичность (форма и ориентация как целевой, так и контрольной полосы вместе), не были информативными для решения задачи (Рис. .1 Б ) (17). Производительность пчел увеличилась за шесть блоков из пяти испытаний (рис.2 A ) (ANOVA для повторных измерений; n = 33 пчелы; эффект блока: F 5, 155 = 18,127, P <0,001). Не было разницы в производительности между группами ниже и выше обученных ( F 1, 31 = 0,422, P = 0,521). Таким образом, данные были объединены и представлены вместе, начиная с Рис. 2 A и далее.Все пчелы, показавшие правильные результаты на ≥ 80% в последних 10 испытаниях этапа 1 (23 пчелы), впоследствии были протестированы с использованием новой мишени для пяти дополнительных испытаний, чтобы определить, могут ли они обобщить задачу на новую цель (стадия 2) (рис. . 1 В ). На этом этапе пчелы выбрали правильную цель в 80 ± 1,9% (среднее ± SEM: n = 23 пчелы) времени, что указывает на то, что они успешно выучили задание (рис. 2 A ). Этот результат значительно отличался от случайного (предполагаемое распределение 50%) с учетом первого выбора (биномиальный тест: P <0.001) или совокупный выбор (один образец т тест: т 21 = 20,120, P <0,001), подтверждающий предыдущие выводы (17).
Мы изменили цели и вертикальное положение целей и контрольных полос псевдослучайно, чтобы исключить возможное использование ассоциативных сигналов. Другими словами, форма, цвет и размер мишеней менялись от испытания к испытанию, а расстояние до мишеней от дна камеры и относительно контрольных стержней варьировалось от испытания к испытанию. Этот формат гарантировал, что низкоуровневые сигналы, такие как совокупная область (область, охватываемая как контрольной полосой, так и целью) или конфигурационная идентичность (форма и ориентация как целевой, так и контрольной полосы вместе), не были информативными для решения задачи (Рис. .1 Б ) (17). Производительность пчел увеличилась за шесть блоков из пяти испытаний (рис.2 A ) (ANOVA для повторных измерений; n = 33 пчелы; эффект блока: F 5, 155 = 18,127, P <0,001). Не было разницы в производительности между группами ниже и выше обученных ( F 1, 31 = 0,422, P = 0,521). Таким образом, данные были объединены и представлены вместе, начиная с Рис. 2 A и далее.Все пчелы, показавшие правильные результаты на ≥ 80% в последних 10 испытаниях этапа 1 (23 пчелы), впоследствии были протестированы с использованием новой мишени для пяти дополнительных испытаний, чтобы определить, могут ли они обобщить задачу на новую цель (стадия 2) (рис. . 1 В ). На этом этапе пчелы выбрали правильную цель в 80 ± 1,9% (среднее ± SEM: n = 23 пчелы) времени, что указывает на то, что они успешно выучили задание (рис. 2 A ). Этот результат значительно отличался от случайного (предполагаемое распределение 50%) с учетом первого выбора (биномиальный тест: P <0.001) или совокупный выбор (один образец т тест: т 21 = 20,120, P <0,001), подтверждающий предыдущие выводы (17).
Экспериментальная установка и конструкция. ( A ) Проектирование камер принятия решений. ( B ) Опытный образец. Этап 1: Ссылка была одинаковой как с левой, так и с правой стороны: горизонтальная черная полоса длиной 5 см и высотой 2 см. Цели были одинаковыми для обеих сторон, но варьировались между испытаниями. На этом этапе использовались пять из шести возможных целей.Награда (2 М сахарозы) или наказание (хинин) помещали в полупрозрачную микроцентрифужную пробирку в центре мишеней (белая трапеция). В ходе испытания пары стимулов были идентичными, за исключением вертикального положения относительно контрольной панели и относительно дна камеры. Между испытаниями цели и положения целей и контрольных полос менялись, так что пчелы могли узнать только отношение целей вверх / вниз к контрольным объектам в качестве предикторов вознаграждения. Примерно половина пчел были вознаграждены по вышеуказанному отношению, а половина — по нижеследующему.Этап 2: тесты передачи не были вознаграждены, и использовалась новая мишень, не использовавшаяся на этапе 1. Этап 3: Сложность варьировалась путем изменения расстояния между мишенью и горизонталью контрольной полосы. Для простых испытаний цель была явно выше или ниже контрольной полосы и не перекрывалась контрольной полосой. Для тяжелых испытаний цели перекрываются с контрольной полосой. Для невозможных испытаний центр обеих мишеней находился на одной линии с контрольной полосой. Этап 4: проверка обобщения правила отказа.Пчелы были обучены изучать новые цели, связанные с наградой и наказанием. Пчелы, прошедшие обучение, были протестированы с запутанными и не получившими вознаграждения целями, объединяющими черты обеих изученных целей.
( B ) Опытный образец. Этап 1: Ссылка была одинаковой как с левой, так и с правой стороны: горизонтальная черная полоса длиной 5 см и высотой 2 см. Цели были одинаковыми для обеих сторон, но варьировались между испытаниями. На этом этапе использовались пять из шести возможных целей.Награда (2 М сахарозы) или наказание (хинин) помещали в полупрозрачную микроцентрифужную пробирку в центре мишеней (белая трапеция). В ходе испытания пары стимулов были идентичными, за исключением вертикального положения относительно контрольной панели и относительно дна камеры. Между испытаниями цели и положения целей и контрольных полос менялись, так что пчелы могли узнать только отношение целей вверх / вниз к контрольным объектам в качестве предикторов вознаграждения. Примерно половина пчел были вознаграждены по вышеуказанному отношению, а половина — по нижеследующему.Этап 2: тесты передачи не были вознаграждены, и использовалась новая мишень, не использовавшаяся на этапе 1. Этап 3: Сложность варьировалась путем изменения расстояния между мишенью и горизонталью контрольной полосы. Для простых испытаний цель была явно выше или ниже контрольной полосы и не перекрывалась контрольной полосой. Для тяжелых испытаний цели перекрываются с контрольной полосой. Для невозможных испытаний центр обеих мишеней находился на одной линии с контрольной полосой. Этап 4: проверка обобщения правила отказа.Пчелы были обучены изучать новые цели, связанные с наградой и наказанием. Пчелы, прошедшие обучение, были протестированы с запутанными и не получившими вознаграждения целями, объединяющими черты обеих изученных целей.
Производительность пчел. ( A ) Процент правильных выборов в зависимости от блоков из пяти испытаний (этап 1) и результатов в тесте передачи без вознаграждения (этап 2). ( B ) Выступление на тестах разной сложности (этап 3). Пчелы лучше показали себя на легком (83,8 ± 2,5%), чем на тяжелом (52.4 ± 2,5%) или невозможных (44,5 ± 7%) испытаний. ( C ) Отключение пропорции относительно easy (горизонтальные полосы) и SE (вертикальные полосы). Пчелы были склонны отказываться от трудных и невозможных испытаний, чем от легких испытаний (стадия 3). Пчелы также значительно чаще отказывались от сложных испытаний (в среднем трудных и невозможных испытаний), чем от легких испытаний. ( D ) Выступление на принудительных и вынужденных испытаниях (этап 3). Пчелы лучше справлялись с жесткими испытаниями без принуждения, чем с испытаниями с применением силы. Пчелы показали лучшие результаты в простых испытаниях (ANOVA для повторных измерений; n = 10; F 1, 9 = 6.028, P <0,036) и выполнялись аналогично для принудительных и не принудительных простых испытаний (ANOVA для повторных измерений; n = 10; F 1, 9 = 0,459, P = 0,515). Этот результат согласуется с тем, что большинство пчел находятся в состоянии низкой неопределенности для большинства простых испытаний. Все столбцы на всех панелях представляют средние значения, а все столбцы ошибок представляют SEM.
Пчелы были склонны отказываться от трудных и невозможных испытаний, чем от легких испытаний (стадия 3). Пчелы также значительно чаще отказывались от сложных испытаний (в среднем трудных и невозможных испытаний), чем от легких испытаний. ( D ) Выступление на принудительных и вынужденных испытаниях (этап 3). Пчелы лучше справлялись с жесткими испытаниями без принуждения, чем с испытаниями с применением силы. Пчелы показали лучшие результаты в простых испытаниях (ANOVA для повторных измерений; n = 10; F 1, 9 = 6.028, P <0,036) и выполнялись аналогично для принудительных и не принудительных простых испытаний (ANOVA для повторных измерений; n = 10; F 1, 9 = 0,459, P = 0,515). Этот результат согласуется с тем, что большинство пчел находятся в состоянии низкой неопределенности для большинства простых испытаний. Все столбцы на всех панелях представляют средние значения, а все столбцы ошибок представляют SEM.
На этапе 3 мы проверили пчел еще в 50 испытаниях различной сложности (рис.1 В ). Для простых испытаний цель была явно выше или ниже контрольной полосы и не перекрывалась с контрольной полосой (этап 3) (рис. 1 B ). Для сложных испытаний цели были смещены, но перекрывались с контрольной полосой (этап 3) (Рис. 1 B ). Для невозможных испытаний (так называемых, потому что объективно они не имели правильного ответа и поэтому были вознаграждены псевдослучайно) центры обеих целей совпадали с центром контрольной полосы (этап 3) (рис. 1 B ).Пчелы лучше справлялись с легкими испытаниями, чем с трудными или невозможными (рис.2 B ) (ANOVA для повторных измерений; n = 16; F 2, 26 = 19,94, P <0,001 ).
Обучение и тестирование на этапах A и B часто занимало более 3 часов, поскольку продолжительность обучения определялась скоростью, с которой пчелы возвращались в устройство из своей колонии. Из 23 пчел, завершивших этапы A и B, только 16 вернулись на этап 3 и только 10 завершили все 50 испытаний этого этапа, на завершение которых обычно уходило еще 4 часа или более.На этапе 3 мишени поочередно представлялись в первой и второй камерах принятия решения (Рис. 1 A ). Когда пчелы были представлены в первой камере принятия решения, у них была возможность отказаться от нее и перелететь во вторую камеру, где находился другой набор целей. Когда пчелы были представлены во второй камере, у них не было возможности отказаться, и их выбор был вынужденным. Мы использовали анализ χ 2 , чтобы сравнить частоту отказа от участия в простых, сложных и невозможных испытаниях. По отдельности, 3 из 10 пчел отказались больше от тяжелых испытаний, чем от легких (рис.3, ряд 6). Независимые значения χ 2 можно суммировать, и суммирование значений для отдельных пчел (19) (рис. 3, строка 6) показало, что как группа эти 10 пчел чаще отказывались от сложных испытаний (трудных и невозможных), чем от простых. испытания (добавка χ 2 ; χ 2 = 25,349, df = 10, P = 0,005). Кроме того, пчелы чаще отказывались от трудных или невозможных испытаний, чем от легких испытаний (рис.2 C ) (добавка χ 2 ; χ 2 = 31,268, df = 20, P = 0.052). Пчелы потенциально могли бы решить эту задачу, либо научившись следить за положением цели относительно эталонов (выше или ниже), либо обращая внимание на геометрию эталонов и целей как на сложные стимулы, но независимо от конкретной стратегии пчел. использовались для решения задачи, так как пчелы в группе больше отказывались от трудных, чем от легких испытаний.
Рис. 3.Сравнение результатов тестов на отдельных пчелах. Верхний ряд: идентификационный номер пчелы. Вторая строка: разница между вероятностью отказа от участия в сложных испытаниях (среднее количество сложных и невозможных) минус вероятность отказа от участия в легких испытаниях.Пчелы упорядочиваются по возрастающей шкале дифференциала отказа. Затенение представляет рейтинг для каждой категории (строки). светлее / белее означает лучшую производительность. Третий ряд: разница в производительности между принудительными и принудительными испытаниями. Четвертая строка: Показатели при передаче первой пробной версии концепции отказа (+, отказался; -, сделал выбор и не отказался). Пятая строка: совокупная эффективность передачи концепции отказа (пять испытаний без вознаграждения). Шестая строка: χ 2 значений вероятности отказа в сложном vs.легкие испытания. Седьмая строка: значений P для каждого теста χ 2 . Рамка с пунктирной линией: показывает лучшие характеристики / измерения в каждом поведенческом тесте. Рамка с пунктирной линией: указывает 7 из 10 пчел, которые отказались больше от трудных испытаний, чем от простых.
Пчелы плохо справлялись с трудными задачами и отказывались от них большего (Рис. 2 B и C ), но улучшило ли их выполнение отказ? Чтобы ответить на этот вопрос, мы сравнили, как часто пчелы правильно решали задачу в жестких испытаниях, когда пчелы могли отказаться (невынужденные испытания), с жесткими испытаниями, когда пчелы не могли отказаться (принудительные испытания).Невыполненные испытания проводились в камере первого решения, потому что пчелы имели доступ к выходному отверстию и, следовательно, могли отказаться от них. Принудительные испытания проводились во второй камере принятия решений, потому что не было выходного порта для использования (Рис. 1 A ). Если пчелы использовали опцию отказа адаптивно и выборочно, то они должны были бы лучше работать в жестких испытаниях без принуждения, чем в принудительных. Если, однако, пчелы чаще отказываются от сложных испытаний, потому что они просто научились избегать этой конфигурации, независимо от достоверности конкретных испытаний, то, по прогнозам, пчелы будут демонстрировать одинаковый уровень производительности в принудительных и невынужденных жестких испытаниях.Объединяя данные по всем 10 пчелам, пчелы лучше справлялись с жесткими испытаниями без принуждения, чем с испытаниями с принудительной нагрузкой (ANOVA для повторных измерений; n = 10; F 1, 9 = 6,028, P = 0,036) ( Рис.2 D ), что указывает на то, что пчелы выбирали выборочно, когда их вероятность успешного решения задачи была самой низкой.
Чтобы оценить, могут ли пчелы перенести концепцию отказа на новую сложную задачу, 10 пчел, завершивших стадию 3, прошли обучение (аналогично стадии 1) для дополнительных 20 испытаний по новой задаче (стадия 4), в которой они пришлось различать две цели (рис.1 В ). После 20 тренировочных испытаний мы протестировали этих 10 пчел с «сбивающими с толку» мишенями без вознаграждения, которые представляли собой комбинацию обеих обученных мишеней (рис. 1 B ) (этап 4). Мы предположили, что, если бы пчелы не были уверены в результате выбора этих новых целей, они бы отказались от решений, которые их содержали. Из 10 пчел, завершивших этапы C и D, ни одна пчела не отказалась от участия во время обучения, что позволяет предположить, что они не выбрали вариант отказа в ответ на новый стимул. Тем не менее, 4 из 10 пчел отказались от участия, когда им были представлены «сбивающие с толку» цели при первом испытании стадии 4 испытания, и по крайней мере три из пяти испытательных испытаний с запутывающими мишенями (рис.3).
Обсуждение
Хотя отдельные пчелы различались по производительности, мы показали, что некоторые пчелы индивидуально и в группе значительно изменили свое поведение выбора в ответ на изменение сложности задачи. Пчелы показали: ( i ) большую вероятность отказа от сложных испытаний по сравнению с легкими испытаниями; ( II ) лучшая производительность в ненасильственных испытаниях по сравнению с жесткими принудительными испытаниями; и ( iii ) перенесен отказ в новую ситуацию.
Производительность сильно различалась среди протестированных пчел (рис.3), но люди, которые значительно чаще отказывались от участия в сложных испытаниях, чем в легких испытаниях, также получили высокие оценки по разнице результатов между принудительными и принудительными испытаниями и тестом обобщения (рис. 3). Это открытие предполагает, что пчелы, которые чаще всего отказываются от тяжелых испытаний, были теми, кто лучше всех справлялся с трудной задачей, а не пчелами, которые не могли различать трудные испытания и избегали их.
Различия в производительности пчел были задокументированы во всех предыдущих исследованиях познания пчел, включая обучение правилам (17, 20), контекстное обучение (21), категоризацию (22) и подсчет (23, 24).Действительно, исследования когнитивных способностей животных в целом сообщают о большой разнородности действий испытуемых (4–9, 11, 25). Поэтому неудивительно, что только некоторые пчелы, подвергшиеся нашему напряженному режиму тестирования, индивидуально продемонстрировали адаптивное использование правила отказа в жестких испытаниях. Эти межиндивидуальные различия могут включать вариации в мотивации, когнитивных способностях, стратегии выбора или любую их комбинацию. Доля пчел, способных обобщить правило отказа на новую задачу в нашем исследовании, была аналогична той, что сообщается для приматов (5).
Наши данные показывают, что некоторые пчелы оказались способными адаптировать свою стратегию принятия решений в ответ на изменение сложности задачи, адаптивно используя возможность отказа. Ранее было показано, что пчелы реагировали на изменение сложности задачи или риска задачи замедлением, чтобы повысить точность (18, 26), а в этологических условиях пчелы реагировали на загадочный риск хищничества изменением своей стратегии добычи пищи, возможно, схоже с выбором из-за трудных различий в нашем анализе (27).Эти данные поднимают вопрос о том, как пчелы это делают. Адаптивное использование возможности отказа в жестких испытаниях интерпретировалось как доказательство того, что животные способны реагировать на степень своей неопределенности (4). Однако утверждалось, что подобные поведенческие результаты можно объяснить простыми ассоциативными механизмами, которые не требуют мониторинга неопределенности. Ле Пелли (14) предполагает, что с помощью механизма ассоциативного обучения силы условных реакций (как положительные, так и отрицательные) будут связаны с различными усвоенными стимулами в задаче различения.Предполагается, что система восприятия зашумлена, поэтому не ожидается, что никакой стимул будет распознаваться идеально, а усвоенные ответы могут быть обобщены на аналогичные стимулы (28). Благодаря обучению сложные стимулы станут ассоциироваться с высокой вероятностью наказания из-за большого количества ошибок в этих испытаниях. Хотя это само по себе не приносит пользы, реакция отказа все же имеет условную силу реакции, поскольку позволяет избежать наказания. Одних этих допущений достаточно, чтобы предсказать, что животные будут отказываться от сложных испытаний больше, чем от легких в определенных парадигмах, и что производительность должна быть лучше в невынужденных, чем в принудительных жестких испытаниях (14, 15).
Таким образом, были предложены два разных механизма для объяснения одного и того же поведенческого феномена, и мы вынуждены отвергнуть более сложный. Однако объективно оценить, какой механизм является наиболее сложным, в настоящее время сложно, потому что, хотя нейробиология ассоциативного обучения хорошо изучена у пчел, у нас нет нейробиологической модели того, как может работать мониторинг неопределенности (если он имеет место). Kepecs et al. (29) утверждают, что оценка неопределенности не более сложна, чем простое распознавание стимула и реакция на него.Если нейронные вычисления соответствия между новым и усвоенным стимулом работают в соответствии с байесовской теорией, то достоверная оценка соответствия будет неотъемлемой частью этого вычисления. У крыс отдельные нейроны орбитофронтальной коры демонстрируют паттерны возбуждения, которые коррелируют со степенью достоверности, моделируемой таким образом (29).
Схема обучения пчел намного проще, чем у любой системы млекопитающих. Благодаря этому открытию медоносные пчелы могут оказаться ценной системой для окончательного определения того, может ли простой мозг животных оценить степень достоверности.У медоносных пчел ассоциативное обучение происходит в цепи, включающей 170 000 клеток Кеньона грибовидных тел и внешних нейронов, которые выходят из грибовидных тел в премоторные области (30–32). Стимулы кодируются в виде разреженного поперечного волокна в популяции клеток Кеньона. Клетки Кеньона и внешние нейроны связаны конвергентной связной матрицей, которая пластична и модифицируется опытом в соответствии с процессами Хебба (33, 34). Обучение включает в себя изменение синаптических весов между клетками Кеньона, активируемыми усвоенным стимулом, и внешними нейронами, запускающими усвоенный ответ (34, 35).В парадигме дифференцированного кондиционирования, в которой пчела обучается тому, что один стимул вознаграждается, а другой нет, во время обучения сенсорные представления двух стимулов расходятся (36, 37). Каждый стимул активирует разные двигательные реакции и различные наборы внешних нейронов (34).
Неоднозначный стимул между двумя усвоенными стимулами потенциально может коактивировать оба возможных моторных ответа. Было высказано предположение, что разрешение этого конфликта, вероятно, достигается взаимным ингибированием между нейронами в популяции внешних нейронов, так что активируется только один доминантный моторный паттерн (33–35).Если эта предполагаемая нейровычислительная структура верна, то степень соответствия между стимулом и условностью выученного стимула-ответа является неотъемлемой частью ассоциативного механизма. Уверенность будет коррелировать со степенью коактивации несовместимых моторных ответов активированными внешними нейронами, степенью, в которой эти конкурирующие возможные выходы могут быть разрешены посредством взаимного торможения, и силой согласованного моторного выхода. Теоретически эта корреляция может работать как нейронный сигнал степени неопределенности, требующий минимальных элементов схемы сверх тех, которые требуются для ассоциативного обучения.Мы не знаем, какие элементы в мозгу пчелы могут позволить оценке неопределенности влиять на вероятность выбора или отказа. У позвоночных нейромодуляторы ацетилхолин и норадреналин участвуют в модуляции выбора посредством неопределенности (38, 39). Таким образом, мы предлагаем, чтобы экспериментальное тестирование того, руководит ли оценка неопределенности поведением пчел, должно быть сосредоточено на вычислениях в матрице ассоциации между клеткой Кеньона и внешними популяциями нейронов мозга пчелы, а также на роли нейромодуляторов в этой системе.
Таким образом, мы показали, что медоносные пчелы способны избирательно избегать выбора, когда информация ограничена, и тем самым повышают свой успех в тесте выбора. Это открытие показывает, что даже простые беспозвоночные способны принимать сложные и адаптивные решения и чувствительны к изменениям сложности заданий. Такое поведение можно объяснить прямой оценкой неопределенности или ассоциативными механизмами, которые по-разному оценивают результаты сложных и легких испытаний.Однако нейронное вычисление неопределенности может быть не более сложным, чем вычисление, необходимое для классификации и ассоциации. Определить, контролируют ли простые беспозвоночные неопределенность в принятии решений — или просто кажется, что они это делают, — можно решить только с помощью электрофизиологического тестирования нейробиологической модели для мониторинга неопределенности. Однако, какая бы вычислительная стратегия не использовалась, наши данные показывают, что способность адаптивно реагировать на трудный выбор может быть более общим, чем считалось ранее.
Материалы и методы
Эксперименты проводились с пчелами, содержащимися в Университете Маккуори (Сидней) в период с декабря 2012 г. по апрель 2013 г. (лето в Южном полушарии). Пчелы принадлежали к стандартному коммерческому поголовью, доступному в Австралии (в основном, из семейства Ligustica), и содержали матку, спарившуюся естественным путем. В наших экспериментах использовались пчелы-фуражиры из семей открытого пчелиного двора, каждая из которых содержала около 30 000 пчел. Пчел-собирателей привлекали к испытательному аппарату раствором сахарозы.Отдельных пчел наносили на грудную клетку краской и приучали из улья летать в камеры принятия решений, расположенные в нескольких метрах от них. Цилиндрические камеры для принятия решений с открытым верхом освещались солнечным светом и закрывались сеткой от насекомых, через которую наблюдатель записывал решения. Вход представлял собой отверстие размером 4 см с внутренними размерами 20 см × 25 см. Внутри каждого цилиндра был белый, отражающий ультрафиолетовый свет фон для целей.
Тренировочные цели на этапах A, B и C (рис.1 B ) представляли собой вертикальную решетку размером 4 × 4 см и радиальный трехсекторный узор размером 4 × 4 см, которые были ахроматическим «черным», а голубо-пурпурно-желто-черный (CMYK) желто-черный (CMYK) размером 4 × 4 см. крест, оранжевый круг CMYK диаметром 4 см, фиолетовый ромб CMYK 5 × 2,5 см и пурпурный прямоугольник CMYK размером 3 × 5 см. Все мишени были напечатаны на лазерном принтере с высоким разрешением. Мишени, питательные трубки, растворы хинина и сахара заменяли между каждым учебным испытанием.
Протокол обучения.
При обучении использовался протокол дифференциальной подготовки.Спаренные цели были размещены по отношению к 2 × 4 см черной прямоугольной опорной панели. Мишень, помещенная над контрольной полосой, содержала 50 мкл 2 М раствора сахарозы. Мишень под контрольной полосой предлагала 50 мкл 50 мМ раствора дигидрата хинина гидрохлорида (который является отталкивающим для пчел). Отдельную группу пчел дрессировали с обратными обстоятельствами. Награда и наказание предлагались в разрезанной прозрачной микроцентрифужной пробирке, прикрепленной к мишеням. В каждом испытании регистрировался первый выбор пчелы.
Пчелы были сначала обучены первой и второй камерам принятия решений с использованием одной из указанных выше целей для двух испытаний, в каждой из которых они получили две правильные конфигурации с вознаграждением. Этот процесс должен был гарантировать, что пчелы знали, что цели могут быть расположены во второй камере. Пчелы не могли повторно войти в первую камеру после того, как они вошли во вторую. На этапе 1 эксперимента (рис. 1 B ), если пчела выбирала правильную цель (например, над контрольной полосой), ей позволяли пить 2 М сахарозы ad libitum.В случае неправильного (например, ниже контрольной полосы), как только они наблюдали за дегустацией раствора хинина хоботком, применяли воздушную затяжку до тех пор, пока пчела не отпустила цель. Испытания чередовались между первой и второй камерой устройства, чтобы пчелы узнали, что полет через первую камеру ведет во вторую камеру принятия решения (рис. 1 A ).
Обобщающий тест.
Во время обобщающих тестов (стадия 4) (рис. 1 B ) пчел тренировали с двумя мишенями, одна награждалась (2 M сахароза), а другая наказывалась (50 мМ хинин + вдох).Целями для обобщающего теста были три алмаза в форме капли, расположенные радиально и окрашенные в HSB (оттенок, насыщенность, яркость: 242, 38, 67) и HSB (54, 56, 89). Испытания чередовались между первой и второй камерой принятия решений, чтобы пчелы знали, что эти цели могут быть расположены внутри любой камеры. Был зафиксирован выбор, когда пчела либо приземлялась на цель, либо пролетала через выходное отверстие (отказ). В любой момент пчела была выпущена, чтобы начать новое испытание. Этот процесс повторялся пять раз на пчелу.
Благодарности
Мы благодарим Мартина Джурфа, Джереми Нивена, Гидона Фельсена и Кена Ченга за комментарии к предыдущим черновикам. C.J.P. был профинансирован стипендией для получения докторской степени Endeavour Awards.
Сноски
Автор: C.J.P. и A.B.B. спланированное исследование; C.J.P. проведенное исследование; C.J.P. и A.B.B. внесены новые реагенты / аналитические инструменты; C.J.P. и A.B.B. проанализированные данные; и C.J.P. и A.B.B. написал газету.
Авторы заявляют об отсутствии конфликта интересов.
Эта статья представляет собой прямое представление PNAS.
(PDF) Распределенная пластичность для обонятельного обучения и памяти в мозге медоносной пчелы Брайан Х. Смит, Рамон Уэрта, Максим Баженов, Ирина Синакевич
407
6.1 Распределенная пластичность для обонятельного обучения и памяти в мозге медоносной пчелы
7. Denker M, Finke R, Schaupp F, Grun S, Menzel R (2010) Нейронные корреляты обучения запаху
в антеннальной доле медоносной пчелы.Eur J Neurosci 31 (1): 119–133
8. Дитцен М., Эверс Дж. Ф., Галиция К. Г. (2003) Сходство запаха не влияет на время, необходимое для обработки запаха
. Chem Senses 28 (9): 781–789
9. Дрезнер-Леви Т., Шафир С. (2007) Параметры переменного распределения вознаграждения, которые влияют на риск
чувствительность медоносных пчел. J Exp Biol 210 (Pt 2): 269–277
10. Фабер Т., Йоргес Дж., Мензель Р. (1999) Ассоциативное обучение изменяет нейронные репрезентации
запахов в мозгу насекомых.Nat Neurosci 2 (1): 74–78
11. Фаруки Т., Робинсон К., Вэссин Х., Смит Б.Х. (2003) Модуляция ранней обонятельной обработки
путем октопаминергического подкрепления у медоносной пчелы. J Neurosci 23: 5370–5380
12. Фернандес П.С., Локателли Ф.Ф., Персона-Реннелл Н., Делео Г., Смит Б.Х. (2009) Ассоциативное условие
настраивает временную динамику ранней обонятельной обработки. J Neurosci 29 (33): 10191–10202
13. Галан Р.Ф., Вейдерт М., Мензель Р., Герц А.В., Галиция К.Г. (2006) Сенсорная память на запахи
кодируется спонтанной коррелированной активностью между обонятельными клубочками.Neural Comput
18 (1): 10–25
14. Галиция К.Г., Ресслер В. (2010) Параллельные обонятельные системы у насекомых: анатомия и функции.
Annu Rev Entomol 55: 399–420
15. Ганешина О., Мензель Р. (2001) ГАМК-иммунореактивные нейроны грибовидных тел
пчелы: электронно-микроскопическое исследование. J Comp Neurol 437 (3): 335–349
16. Джурфа М. (2007) Поведенческий и нейронный анализ ассоциативного обучения медоносной пчелы: вкус
из волшебного колодца.J Comp Physiol A 193 (8): 801–824
17. Guerrieri F, Schubert M, Sandoz JC, Giurfa M (2005) Перцептивное и нейронное обонятельное сходство
у медоносных пчел. PLoS Biol 3 (4): e60
18. Hammer M (1993) Идентифицированный нейрон опосредует безусловный стимул в ассоциативном обонятельном обучении
медоносных пчел. Nature 366: 59–63
19. Hildebrand JG, Shepherd GM (1997) Механизмы обонятельной дискриминации: сходящиеся
свидетельства общих принципов для разных типов.Annu Rev Neurosci 20: 595–631
20. Hourcade B, Perisse E, Devaud JM, Sandoz JC (2009) Долговременная память формирует первичный обонятельный центр
мозга насекомого. Изучите Mem 16 (10): 607–615
21. Huerta R, Nowotny T (2009) Быстрое и надежное обучение с помощью сигналов подкрепления: исследования
в мозге насекомого. Neural Comput 21 (8): 2123–2151
22. Joerges J, Kuttnet A, Galizia G, Menzel R (1997) Представления запахов и запахов
смесей, визуализированных в мозгу медоносной пчелы.Nature 387: 285–288
23. Kreissl S, Eichmüller S, Bicker G, Rapus J, Eckert M (1994) Octopamine-like immunoreac-
активность в головном мозге и субэзофагеальных ганглиях медоносной пчелы. J Comp Neurol
348 (4): 583–595
24. Laurent G (2002) Динамика обонятельной сети и кодирование многомерных сигналов.
Nat Rev Neurosci 3 (11): 884–895
25. LeCun Y, Cortes Chylcem (1998) База данных MNIST.
http: // yannlecuncom / exdb / mnist /
26.Linster C, Menon AV, Singh CY, Wilson DA (2009) Специфическое для запаха привыкание возникает из
взаимодействия афферентной синаптической адаптации и внутренней синаптической потенциации в обонятельной
коре головного мозга. Изучите Mem 16 (7): 452–459
27. Lubow RE (1973) Скрытое торможение. Psychol Bull 79: 398–407
28. Марр Д. (1969) Теория коры мозжечка. J Physiol 202 (2): 437–470
29. Mauelshagen J (1993) Нейронные корреляты парадигм обонятельного обучения в идентифицированном нейроне
в мозге медоносной пчелы.J Neurophysiol 69 (2): 609–625
30. Menzel R (1990) Обучение, память и «познание» у медоносных пчел. В: Кеснер Р.П., Олтон DS
(ред.) Нейробиология сравнительного познания. Лоуренс Эрлбаум, Хиллсдейл, стр. 237–292
31. Мензель Р., Манц Г. (2005) Нейронная пластичность внешних нейронов грибовидного тела в мозге пчелы-пчелы
. J Exp Biol 208 (Pt 22): 4317–4332
32. Okada R, Rybak J, Manz G, Menzel R (2007) Связанная с обучением пластичность PE1 и других
внешних нейронов грибовидного тела в мозге медоносной пчелы.J Neurosci 27 (43): 11736–11747
33. Перес-Орив Дж., Мазор О., Тернер Г.К., Кассенаер С., Уилсон Р.И. и др. (2002) Осцилляции и
редких представлений запаха в теле гриба. Science 297 (5580): 359–365
Расшифровка спектральных профилей мощности по активности мозга FMRI во время естественного слухового опыта
Абрамс, Д.А., Ряли, С., Чен, Т., Хордиа, П., Хузам, А. , Левитин, DJ, и Менон, В. (2013). Межпредметная синхронизация реакций мозга при прослушивании естественной музыки. Европейский журнал нейробиологии , 37 (9), 1458–1469.
Артикул PubMed PubMed Central Google ученый
Аллури, В., Тойвиайнен, П., Яаскелайнен, И. П., Глереан, Э., Самс, М., & Браттико, Э. (2012). Крупномасштабные мозговые сети возникают в результате динамической обработки музыкального тембра, тональности и ритма. NeuroImage , 59 (4), 3677–3689.
Артикул PubMed Google ученый
Аллури, В., Тойвиайнен, П., Лунд, Т. Е., Валлентин, М., Вууст, П., Нанди, А. К., Ристаниеми, Т., и Браттико, Э. (2013). От Вивальди до Битлз и обратно: прогнозирование латерализованной реакции мозга на музыку. NeuroImage , 83 , 627–636.
Артикул PubMed Google ученый
Angenstein, N., & Brechmann, A. (2015). Обработка звуковой интенсивности: категоризация против сравнения. NeuroImage , 119 , 362–370.
Артикул PubMed Google ученый
Bartels, A., & Zeki, S. (2005). Динамика мозга в естественных условиях просмотра — новое руководство по отображению связи in vivo. NeuroImage , 24 (2), 339–349.
Артикул PubMed Google ученый
Bilecen D, Seifritz E, Scheffler K, Henning J, AC S (2002) Амплитопичность слуховой коры человека: исследование фМРТ.NeuroImage 17 (2): 710–718.
Bordier, C., Puja, F., & Macaluso, E. (2013). Сенсорная обработка при просмотре кинематографического материала: компьютерное моделирование и функциональная нейровизуализация. NeuroImage , 67 , 213–226.
Артикул PubMed Google ученый
Конг, Ф., Аллури, В., Нанди, А. К., Тойвиайнен, П., Руи, Ф., Абу-Джамус, Б., Гонг, Л., Кренен, Б. Г. У., Пойконен, Х., & Хуотилайнен, М. (2013). Связывание реакций мозга с натуралистической музыкой посредством анализа текущих характеристик ЭЭГ и стимулов. Мультимедийные транзакции IEEE на , 15, (5), 1060–1069.
Артикул Google ученый
Кортес, К., и Вапник, В. (1995). Сети опорных векторов. Машинное обучение , 20 (3), 273–297.
Google ученый
Дыкстра, А.Р., Кох, К. К., Брейда, Л. Д., и Марк Джуд, Т. (2012). Диссоциация обнаружения и различения чистых тонов после двусторонних поражений слуховой коры. PloS One , 7 (9), e44602.
CAS Статья PubMed PubMed Central Google ученый
Фанг, Дж., Ху, X., Хан, Дж., Цзян, X., Чжу, Д., Го, Л., и Лю, Т. (2015). Управляемый данными анализ функциональных взаимодействий мозга при свободном прослушивании музыки и речи. Визуализация мозга и поведение , 9 (2), 162–177.
Артикул PubMed Google ученый
Фарбуд, М. М., Хигер, Д. Дж., Маркус, Г., Хассон, Ю., и Лернер, Ю. (2015). Нейронная обработка иерархической структуры в музыке и речи в разных временных масштабах. Frontiers in Neuroscience , 9 , 157.
Статья PubMed PubMed Central Google ученый
Фрей, Б.Дж. И Д. Дук (2007). Кластеризация путем передачи сообщений между точками данных. Наука , 315 (5814), 972–976.
CAS Статья PubMed Google ученый
Хан, Дж., Чен, К., Шао, Л., Ху, X., Хан, Дж., И Лю, Т. (2015). Изучение вычислительных моделей запоминаемости видео на основе изображений мозга с помощью FMRI. IEEE Trans. По кибернетике , 45 (8), 1692–1703.
Артикул Google ученый
Ханке, М., Баумгартнер, Ф. Дж., Ибе, П., Кауле, Ф. Р., Поллманн, С., Спек, О., Зинке, В., и Штадлер, Дж. (2014). Набор данных фМРТ с высоким разрешением 7 тесла от сложной естественной стимуляции с аудио-фильмом. Scientific Data , 1 , 140003.
Статья PubMed PubMed Central Google ученый
Хассон, У., и Хани, К. (2012). Будущие тенденции в нейровизуализации: нейронные процессы, выраженные в контексте реальной жизни. NeuroImage , 62 (2), 1272–1278.
Артикул PubMed PubMed Central Google ученый
Hasson, U., Nir, Y., Levy, I., Fuhrmann, G., & Malach, R. (2004). Межпредметная синхронизация корковой активности при естественном зрении. Наука , 303 (5664), 1634–1640.
CAS Статья PubMed Google ученый
Хинтон, Г.Э. (2002). Обучающие продукты экспертов за счет минимизации контрастных расхождений. Нейронные вычисления , 14 (8), 1771–1800.
Артикул PubMed Google ученый
Ху, X., Lv, C., Cheng, G., Lv, J., Guo, L., Han, J., & Liu, T. (2015). ФМРТ-декодирование с ограничением разреженности визуальной заметности в натуралистических видеопотоках. Автономное умственное развитие, транзакции IEEE по телефону 7 , 2 , 65–75.
Google ученый
Хут, А. Г., Нисимото, С., Ву, А. Т., и Галлант, Дж. Л. (2012). Непрерывное семантическое пространство описывает представление тысяч категорий объектов и действий в человеческом мозгу. Нейрон , 76 (6), 1210–1224.
CAS Статья PubMed PubMed Central Google ученый
Дженкинсон, М., Бекманн, К. Ф., Беренс, Т. Е., Вулрич, М. В., и Смит, С. М. (2012). FSL. NeuroImage , 62 (2), 782–790.
Артикул PubMed Google ученый
Цзи, X., Хан, Дж., Цзян, X., Ху, X., Го, Л., Хан, Дж., Шао, Л., и Лю, Т. (2015). Анализ музыки / речи посредством интеграции аудиоконтента и функциональной реакции мозга. Информационные науки , 297 , 271–282.
Артикул Google ученый
Кауппи, Дж.П., Паюла Дж. И Тока Дж. (2014). Универсальный программный пакет для межпредметного корреляционного анализа фМРТ. Границы нейроинформатики , 8 , 2.
Статья PubMed PubMed Central Google ученый
Кляйн, М. Э., и Заторре, Р. Дж. (2015). Представления инвариантных музыкальных категорий можно декодировать с помощью анализа паттернов локально распределенных жирных ответов в верхних височных и внутри теменных бороздах. Кора головного мозга , 25 (7), 1947–1957.
Артикул PubMed Google ученый
Кумар, С., Бонничи, Х. М., Теки, С., Агус, Т. Р., Пресснитцер, Д., Магуайр, Э. А., и Т. Д., Г. (2014). Представления определенных акустических паттернов в слуховой коре и гиппокампе. Proceedings Biological Sciences / The Royal Society , 281 (1791), 20141000.
Статья Google ученый
Лангерс, Д.Р., Ван, Д. П., Шенмейкер, Э. С., и Бэкес, В. Х. (2007). Активация фМРТ в зависимости от интенсивности и громкости звука. NeuroImage , 35 (2), 709–718.
Артикул PubMed Google ученый
Lasota, K., Ulmer, J., Firszt, J., Biswal, B., Daniels, D., & Prost, R. (2003). Зависимая от интенсивности активация первичной слуховой коры при функциональной магнитно-резонансной томографии. Журнал компьютерной томографии , 27 (2), 213–218.
Артикул PubMed Google ученый
Локвуд, А., Салви, Р., Мл, А.С., Вак, Д., Мерфи, Б., и Буркард, Р. (1999). Функциональная анатомия нормальной слуховой системы человека: ответы на тоны 0,5 и 4,0 кГц различной интенсивности. Кора головного мозга , 9 (1), 65–76.
CAS Статья PubMed Google ученый
Мор, К.М., Кинг, В. М., Фриман, А. Дж., Бриггс, Р. В., и Леонард, К. М. (1999). Влияние интенсивности речевых стимулов на активацию слуховой коры исследовали с помощью функциональной магнитно-резонансной томографии. Журнал Акустического общества Америки , 105 (5), 2738–2745.
CAS Статья Google ученый
Mustovic, H., Scheffler, K., Di Salle, F., Esposito, F., Neuhoff, J.G., Hennig, J., & Seifritz, E.(2003). Временная интеграция последовательных слуховых событий: период молчания в звуковой структуре активирует человеческое тело. NeuroImage , 20 (1), 429–434.
Артикул PubMed Google ученый
Нардо, Д., Сантанджело, В., и Макалузо, Э. (2011). Стимул-ориентированная ориентация зрительно-пространственного внимания в сложных динамических средах. Нейрон , 69 (5), 1015–1028.
CAS Статья PubMed Google ученый
Населарис Т., Кей, К. Н., Нисимото, С., и Галлант, Дж. Л. (2011). Кодирование и декодирование в фМРТ. NeuroImage , 56 (2), 400–410.
Артикул PubMed Google ученый
Нисимото, С., Ву, А. Т., Населарис, Т., Бенджамини, Ю., Ю, Б., и Галлант, Дж. Л. (2011). Реконструкция визуальных впечатлений от мозговой активности, вызванной естественными фильмами. Текущая биология , 21 (19), 1641–1646.
CAS Статья PubMed PubMed Central Google ученый
Норман, К.А., Полин, С. М., Детре, Г. Дж., И Хаксби, Дж. В. (2006). Помимо чтения мыслей: анализ множественных вокселей данных фМРТ. Тенденции в когнитивных науках , 10 (9), 424–430.
Артикул PubMed Google ученый
Опиц, Б., Ринне, Т., Меклингер, А., Фон Крамон, Д. Ю., и Шрегер, Э. (2002). Дифференциальный вклад лобной и височной коры в обнаружение слуховых изменений: результаты фМРТ и ERP. NeuroImage , 15 (1), 167–174.
Артикул PubMed Google ученый
Проакис, Дж. Г., и Манолакис, Д. Г. (1992). Цифровая обработка сигналов: принципы, алгоритмы и приложения . Максвелл Макмиллан Канада, Максвелл Макмиллан Интернешнл: Macmillan.
Google ученый
Рейтерер, С., Эрб, М., Гродд, В., И Вильдгрубер Д. (2008). Церебральная обработка тембра и громкости: данные фМРТ о вкладе области Брока в базовую слуховую дискриминацию. Визуализация мозга и поведение , 2 (1), 1–10.
Артикул Google ученый
Рель, М., & Уппенкамп, С. (2012). Нейронное кодирование интенсивности и громкости звука в слуховой системе человека. Джаро , 13 (3), 369–379.
Артикул PubMed PubMed Central Google ученый
Саенс, М., & Лангерс, Д. (2014). Тонотопическое картирование слуховой коры человека. Исследование слуха , 307 (1), 42–52.
Артикул PubMed Google ученый
Санторо, Р., Мурель, М., Де, М. Ф., Гебель, Р., Угурбил, К., Якуб, Э., и Формизано, Э. (2014). Кодирование естественных звуков с множеством спектральных и временных разрешений в слуховой коре человека. PLoS Computational Biology , 10 (1), e1003412.
Артикул PubMed PubMed Central Google ученый
Спайерс, Х. Дж., И Магуайр, Э. А. (2007). Расшифровка активности человеческого мозга во время реальных событий. Тенденции в когнитивных науках , 11 (8), 356–365.
Артикул PubMed Google ученый
Talavage, T. M., Sereno, M. I., Melcher, J. R., Ledden, P.J., Rosen, B.Р. и Дейл А. М. (2004). Тонотопическая организация в слуховой коре человека, выявленная прогрессией частотной чувствительности. Журнал нейрофизиологии , 91 (3), 1282–1296.
Артикул PubMed Google ученый
Тойвиайнен, П., Аллури, В., Браттико, Э., Валлентин, М., и Вууст, П. (2013). Захват музыкального мозга с помощью лассо: динамическое декодирование музыкальных элементов из данных фМРТ. NeuroImage , 88C , 170–180.
Google ученый
Trost, W., Frühholz, S., Cochrane, T., Cojan, Y., & Vuilleumier, P. (2015). Временная динамика музыкальных эмоций исследуется через межпредметную синхронизацию мозговой активности. Социальная когнитивная и аффективная нейробиология. DOI: 10.1093 / сканирование / nsv060.
PubMed PubMed Central Google ученый
Уппенкамп, С., & Рель, М.(2013). Слуховая нейровизуализация человека по интенсивности и громкости. Исследование слуха , 307 (1), 65–73.
PubMed Google ученый
Велч П. Д. (1967). Использование быстрого преобразования Фурье для оценки спектров мощности: метод, основанный на усреднении по времени по коротким модифицированным периодограммам. Транзакции IEEE по аудио и электроакустике , 15 (2), 70–73.
Артикул Google ученый
Zhao S, Jiang X, Han J, Hu X, Zhu D, Lv J, Zhang T, Guo L, Liu T (2014) Расшифровка слуховой заметности с помощью FMRI изображений мозга.Статья представлена в материалах международной конференции ACM по мультимедиа, Орландо, Флорида, США.
От молекул к обществу: механизмы, регулирующие роение у медоносных пчел (Apis spp.)
Alaux, C., Duong, N., Schneider, SS, Southey, BR, Rodriguez-Zas, S., Robinson, GE (2009) Качество модулирующего сигнала связи связано с определенным нейрогеномным состоянием у медоносных пчел. PLoS ONE 4 (8), e6694
PubMed PubMed Central Статья CAS Google ученый
Аллен, М.Д. (1955) Наблюдения за пчелами, сопровождающими свою матку. Anim. Behav. 3 , 66–69
Артикул Google ученый
Аллен, доктор медицины (1956) Поведение пчел, готовящихся к рою. Anim. Behav. 4 (1), 14–22
Артикул Google ученый
Allen, M.D. (1959) Возникновение и возможное значение «встряхивания» пчелиных маток рабочими.Anim. Behav. 7 , 66–69
Артикул Google ученый
Амвросий, Дж. Т. (1976) Транзитные рои. Пчелиный мир 57 , 101–109
Google ученый
Анхольт Р.Р., Маккей Т.Ф. (2012) Генетика агрессии. Анну. Преподобный Жене. 46 , 145–164
PubMed CAS Статья Google ученый
Авитабиле, А., Морс, Р.А., Бох, Р. (1975) Рой медоносных пчел, руководствуясь феромонами. Аня. Энтомол. Soc. Являюсь. 68 , 1079–1082
Google ученый
Бикман, М., Фатке, Р.Л., Сили, Т.Д. (2006) Как информированное меньшинство разведчиков направляет рой медоносных пчел в свой новый дом? Anim. Behav. 71 , 161–171
Артикул Google ученый
Бен-Шахар, Ю., Робишон, А., Соколовски, М.Б., Робинсон, Г.Е. (2002) Влияние действия генов в разных временных масштабах на поведение. Наука 296 (5568), 741–744
PubMed CAS Статья Google ученый
Бернаскони, Г., Ратниекс, F.L.W., Рэнд, Э. (2000) Влияние «опрыскивания» при борьбе с матками медоносных пчел ( Apis mellifera L.) на временную структуру схваток. Insectes Soc. 47 , 21–26
Артикул Google ученый
Бисмейер, Дж.C. (2003) Возникновение и контекст сигнала тряски у медоносных пчел ( Apis mellifera ), использующих естественные источники пищи. Этология 109 (12), 1009–1020
Статья Google ученый
Блюм, М.С. (1992) Медоносные пчелиные феромоны. В: Грэм, Дж. М. (ред.) Улей и медоносная пчела, стр. 269–361. Дадант и сыновья, Гамильтон
Google ученый
Бох, Р., Ленский Ю. (1976) Феромонный контроль выращивания маток в семьях медоносных пчел. J. Apic. Res. 15 , 59–62
CAS Google ученый
Boch, R., Shearer, D.A., Young, J.C. (1975) Феромоны медоносных пчел: полевые испытания натурального и искусственного вещества матки. J. Chem. Ecol. 1 (1), 133–148
CAS Статья Google ученый
Порода, М.Д., Стиллер Т.М., Блюм М.С., Пейдж Р.Э. (1992) Распознавание гнездовых пчел: эффекты фекальных феромонов матки. J. Chem. Ecol. 18 , 1633–1640
PubMed CAS Статья Google ученый
Bruinsma, O., Van Kruijt, J.P., Dusseldorp, W. (1981) Задержка появления маток медоносной пчелы Apis mellifera в ответ на звук гудения. Proc. К. Нед. Акад. van Wet. Сер. C Biol. Med. Sci. 84 , 381–387
Google ученый
Батлер, К.Г. (1940) Возраст пчел в стае. Пчелиный мир 21 , 9–10
Google ученый
Батлер, К.Г. (1960) Значение вещества матки в роении и вытеснении в колониях медоносных пчел ( Apis mellifera L.). Proc. R. Entomol. Soc. Лондон A 35 , 129–132
Google ученый
Батлер, К.Г., Симпсон, Дж. (1967) Феромоны пчелиной матки ( Apis mellifera L.), которые позволяют ее рабочим следовать за ней во время роения. Proc. R. Entomol. Soc. A 42 (10–12), 149–154
CAS Google ученый
Батлер, К.Г., Каллоу, Р.К., Чапман, Дж. Р. (1964) 9-гидроксидек- транс -2-еновая кислота, феромон, стабилизирующий стаи медоносных пчел. Природа 201 (4920), 733
CAS Статья Google ученый
Батлер, К.G., Fletcher, D.J.C., Watler, D. (1969) Маркировка входа в гнездо феромонами медоносной пчелой Apis mellifera L. и осой Vespula vulgaris L. Anim. Behav. 17 (1), 142–147
Артикул Google ученый
Cao, TT, Hyland, KM, Malechuck, A., Lewis, LA, Schneider, SS (2007) Влияние вибрационного сигнала на взаимодействие рабочих с гнездом и товарищами по гнезду в созданных и недавно основанных колониях медоносная пчела, Apis mellifera .Insectes Soc. 54 , 144–149
Артикул Google ученый
Cao, TT, Hyland, KM, Malechuk, A., Lewis, LA, Schneider, SS (2009) Влияние повторяющихся вибрационных сигналов на поведение рабочих в созданных и недавно основанных колониях медоносных пчел, Apis mellifera . Behav. Ecol. Sociobiol. 63 , 521–529
Артикул Google ученый
Клейтон, Д.F. (2004) Геномика певчих птиц: методы, механизмы, возможности и подводные камни. Аня. Акад. Sci. 1016 , 45–60
PubMed CAS Статья Google ученый
Комбс-младший, Г.Ф. (1972) Набухание стаи рабочих пчел. J. Apic. Res. 11 (3), 121–128
Google ученый
Кузин И.Д., Краузе Дж., Фрэнкс Н.Р., Левин С.А. (2005) Эффективное руководство и принятие решений в группах животных на ходу. Природа 433 , 513–516
PubMed CAS Статья Google ученый
Каммингс М.Э. (2012) Поиск полового отбора в женском мозгу. Филос. Пер. R. Soc. Лондон. B Biol. Sci. 367 (1600), 2348–2356
PubMed PubMed Central Статья Google ученый
Денлингер, Д.Л. (1986) Покой тропических насекомых. Анну. Преподобный Энтомол. 31 , 239–264
PubMed CAS Статья Google ученый
Denlinger, D.L. (2002) Регулирование диапаузы. Анну. Преподобный Энтомол. 47 , 93–122
PubMed CAS Статья Google ученый
Донахью, К., Льюис, Л.А., Шнайдер, С.С. (2003) Роль сигнала вибрации в процессе охоты за домом пчелиного роя ( Apis mellifera ).Behav. Ecol. Sociobiol. 54 , 593–600
Артикул Google ученый
Дуангпхакди О., Хепбур Х. Insectes Soc. 59 , 571–577
Артикул Google ученый
Dyer, F.C. (2002) Биология танцевального языка.Анну. Преподобный Энтомол. 47 , 917–949
PubMed CAS Статья Google ученый
Dyer, F.C., Seeley, T.C. (1994) Миграция колоний тропической медоносной пчелы Apis dorsata F. (Hymenoptera: Apidae). Insectes Soc. 41 , 129–140
Артикул Google ученый
Энгельс, В., Адлер, А., Розенкранц, П., Любке, Г., Франк, В.(1993) Дозозависимое ингибирование экстренного выращивания маток синтетическим 9-ODA у медоносной пчелы, Apis mellifera carnica . J. Comp. Physiol. B. 163 , 363–366
CAS Статья Google ученый
Эш, Х. (1967) Звук, издаваемый роением медоносных пчел. Z. Vergl. Physiol. 56 , 408–411
Артикул Google ученый
Эш, Х.(1976) Температура тела и летные характеристики медоносных пчел в аэродинамической трубе с сервомеханическим управлением. J. Comp. Physiol. 109 , 264–277
Артикул Google ученый
Фефферман, Н.Х., Старкс, П.Т. (2006) Подход к моделированию роения медоносных пчел ( Apis mellifera ). Insectes Soc. 53 (1), 37–45
Артикул Google ученый
Фергюсон, А.W., Free, J.B., Pickett, J.A., Winder, M. (1979) Методы изучения феромонов медоносных пчел, участвующих в кластеризации, и эксперименты по влиянию феромонов Насонова и королевы. Physiol. Энтомол. 4 , 339–344
CAS Статья Google ученый
Fletcher, D.J.C. (1978a) Влияние вибрационных танцев рабочих пчел на деятельность маток-девственниц. J. Apic. Res. 17 , 3–13
Google ученый
Флетчер Д.J.C. (1978b) Вибрация маточников рабочими пчелами и ее связь с проблемой стаи с девственными матками. J. Apic. Res. 17 , 14–26
Google ученый
Fluri, P., Luscher, M., Wille, H., Gerig, L. (1982) Изменения веса глоточной железы и гемолимфы титров ювенильного гормона, белка и вителлогенина у рабочих медоносных пчел. J. Insect Physiol. 28 , 61–68
CAS Статья Google ученый
Фри, Дж.Б. (1987) Феромоны социальных пчел. Издательство Корнельского университета, Итака
Google ученый
Free, J.B., Ferguson, A.W., Pickett, J.A. (1981a) Оценка различных компонентов феромона Насанова, используемых при группировании медоносных пчел. Physiol. Энтомол. 6 (3), 263–268
CAS Статья Google ученый
Фри, Дж. Б., Пикетт, Дж. А., Фергюсон, А.W., Smith, M.C. (1981b) Синтетические феромоны для привлечения роя медоносных пчел ( Apis mellifera ). J. Agr. Sci. 97 (2), 427–431
CAS Статья Google ученый
Гал, Р.А. (1975) Танец пчелоносцев: свидетельство дискриминации по возрасту. Anim. Behav. 23 , 230–232
Артикул Google ученый
Гэри, Н.E. (1962) Аттрактанты химического спаривания у пчелиной матки. Наука 136 , 773–774
PubMed CAS Статья Google ученый
Getz, W.M., Brückner, D., Parisian, T.R. (1982) Родовая структура и роение медоносной пчелы Apis mellifera . Behav. Ecol. Sociobiol. 10 , 265–270
Артикул Google ученый
Гилли Д.C. (1998) Идентичность разведчиков гнезд в стае медоносных пчел. Apidologie 29 (3), 229–240
Статья Google ученый
Гилли, округ Колумбия (2001) Поведение медоносных пчел ( Apis mellifera ligustica ) во время дуэлей маток. Этология 107 , 601–622
Статья Google ученый
Гилли, округ Колумбия (2003) Отсутствие кумовства в преследовании маток на дуэлях пчеловодами.Proc. R. Soc. Лондон. B 270 (1528), 2045–2049
Артикул Google ученый
Гилли, округ Колумбия, Тарпи, Д. (2005) Три механизма уничтожения маток в роящихся колониях медоносных пчел. Apidologie 36 , 461–474
Статья Google ученый
Gilley, D.C., DeGrandi-Hoffman, G., Hooper, J.E. (2006) Летучие соединения, выделяемые живыми матками европейской медоносной пчелы ( Apis mellifera ).J. Insect Physiol. 2 , 520–527
Артикул CAS Google ученый
Grooters, H.J. (1987) Влияние трубок маток и поведения рабочих на время появления маток медоносных пчел. Insectes Soc. 34 , 181–193
Артикул Google ученый
Grozinger, C.M. (в печати) Феромоны пчелиного меда. В: Дж. Грэм (редактор) Улей и медоносная пчела, Индианаполис, Дадан.42 p.
Grozinger, C.M., Robinson, G.E. (2007) Эндокринная модуляция гена, отвечающего за феромон, в мозге медоносной пчелы. J. Comp. Bio. A 193 (4), 461–470
CAS Google ученый
Grozinger, C.M., Sharabash, N.M., Whitfield, C.W., Robinson, G.E. (2003) Феромон-опосредованная экспрессия гена в мозге медоносной пчелы. Proc. Natl. Акад. Sci. США 100 (Дополнение 2), 14519–14525
PubMed CAS PubMed Central Статья Google ученый
Grozinger, C.М., Фан, Ю., Гувер, С.Э., Уинстон, М.Л. (2007a) Полногеномный анализ показывает различия в паттернах экспрессии генов мозга, связанных с кастовой принадлежностью и репродуктивным статусом медоносных пчел ( Apis mellifera ). Мол. Ecol. 16 (22), 4837–4848
PubMed CAS Статья Google ученый
Grozinger, C.M., Fischer, P., Hampton, J.E. (2007b) Разъединение реакции праймера и высвобождающего агента на феромон у медоносных пчел.Naturwissenschaften 94 (5), 375–379
PubMed CAS Статья Google ученый
Хан, Д.А., Денлингер, Д.Л. (2011) Энергетика диапаузы насекомых. Анну. Преподобный Энтомол. 56 , 103–121
PubMed CAS Статья Google ученый
Hefetz, A., Taghizadeh, T., Francke, W. (1996) Экзокринология шмелей-маток Bombus terrestris (Hymenoptera: Apidae, Bombini).Z. Naturforsch. 51 , 409–422
CAS Google ученый
Генрих Б. (1979) Терморегуляция африканских и европейских медоносных пчел во время кормодобывания, нападения и выхода и возвращения из улья. J. Exp. Bio. 80 , 217–229
Google ученый
Генрих Б. (1981) Механизмы и энергетика регулирования температуры пчелиного роя. J. Exp.Биол. 91 , 25–55
Google ученый
Хепберн, Х.Р. (2006) Бегство, миграция и роение медоносных пчел: экологическая и эволюционная перспектива. В кн .: Кипятков В. (ред.) Жизненные циклы социальных насекомых: поведение, экология и эволюция, стр. 121–135. Издательство СПбГУ, Санкт-Петербург
Google ученый
Хепберн, Х.Р., Радлофф, С.E. (1998) Медоносные пчелы Африки, стр. 386. Springer, Нью-Йорк
Google ученый
Гувер, S.E.R., Килинг, C.I., Уинстон, М.Л., Слессор, К.Н. (2003) Влияние феромонов матки на развитие яичников медоносных пчел. Naturwissenschaften 90 , 477–480
PubMed CAS Статья Google ученый
Хуанг, З.Й., Робинсон, Г.Э. (1995) Сезонные изменения титров ювенильных гормонов и скорости биосинтеза у медоносных пчел.J. Comp. Physiol. Б. 165 , 18–28
PubMed CAS Статья Google ученый
Хуанг, З.Й., Робинсон, Г.Э. (1996) Регулирование разделения труда медоносных пчел по возрастной демографии колонии. Behav. Ecol. социол. 39 (3), 147–158
Google ученый
Иноуэ, Т., Сакагами, С., Салмах, С., Нукмал, Н. (1984) Обнаружение успешного бегства безжальной пчелы Trigona (Tetragonula) laeviceps .J. Apic. Res. 23 , 136–142
Google ученый
Янсон, С., Миддендорф, М., Бикман, М. (2005) Стаи медоносных пчел: как разведчики направляют рой неосведомленных пчел? Anim. Behav. 70 , 349–358
Артикул Google ученый
Kankare, M., Salminen, T., Laiho, A., Vesala, L., Hoikkla, A. (2010) Изменения в экспрессии генов, связанные с репродуктивной диапаузой взрослых особей у северных солодовых мух: ген-кандидат микрочиповое исследование.BMC Ecol. 10 , 3
PubMed PubMed Central Статья CAS Google ученый
Кацав-Гозанский, Т., Сорокер, В., Франке, В., Хефец, А. (2003) Пчелы-яйцекладчики имитируют сигнал матки. Insectes Soc. 50 , 20–23
Артикул Google ученый
Килинг, К.И., Слессор, К.Н., Хиго, Х.А., Уинстон, М.Л. (2003) Новые компоненты медоносной пчелы ( Apis mellifera L.) феромон королевской свиты. P. Natl. Акад. Sci.USA 100 (8), 4486–4491
CAS Статья Google ученый
Кочер, С.Д., Грозингер, К.М. (2011) Сотрудничество, конфликт и эволюция феромонов королевы. J. Chem. Ecol. 37 (11), 1263–1275
PubMed CAS Статья Google ученый
Кенигер, Н., Кенигер, Г. (1980) Наблюдения и эксперименты по миграции и танцевальной коммуникации Apis dorsata в Шри-Ланке.J. Apic. Res. 19 , 21–34
Google ученый
Kryger, P., Moritz, R.F.A. (1997) Отсутствие распознавания родства у роящихся пчел ( Apis mellifera ). Behav. Эко. Sociobiol. 40 (4), 271–276
Артикул Google ученый
Kunert, K., Crailsheim, K. (1988) Сезонные изменения содержания углеводов, липидов и белков у молодых рабочих медоносных пчел и их смертность.J. Apic. Res. 27 , 13–21
CAS Google ученый
Ленский, Ю., Слабецкий, Ю. (1981) Ингибирующее действие феромона следа пчелиной матки ( Apis mellifera L.) на построение чашечек роящихся маток. J. Insect Physiol. 27 (5), 313–323
Артикул Google ученый
Лета, М.А., Гилберт, К., Морс, Р.A. (1996) Уровни сахара гемолимфы и гликогена тела медоносных пчел ( Apis mellifera L.) из колоний, готовящихся к рою. J. Insect Physiol. 42 (3), 239–245
CAS Статья Google ученый
Льюис, Л.А., Шнайдер, С.С. (2000) Модуляция поведения рабочего с помощью сигнала вибрации во время охоты за домом в стаях медоносных пчел, Apis mellifera . Behav. Ecol. Sociobiol. 48 , 154–164
Артикул Google ученый
Льюис, Л.А., Шнайдер, С.С. (2008) Танцы миграции в роящихся колониях медоносной пчелы Apis mellifera . Apidologie 39 , 354–361
Статья Google ученый
Ли С.И., Пуругганан М.Д. (2011) Кооперативная амеба: Dictyostelium как модель социальной эволюции. Тенденции Genet. 27 (2), 48–54
PubMed Статья CAS Google ученый
Лян, З.С., Нгуен, Т., Маттила, Х.Р., Родригес-Зас, С.Л., Сили, Т.Д., Робинсон, Г.Э. (2012) Молекулярные детерминанты разведывательного поведения медоносных пчел. Наука 335 , 1225–1228
PubMed CAS Статья Google ученый
Линдауэр, М. (1955) Schwarmbienen auf Wohnungssuche. Z. vergl. Physiol. 37 , 263–324
Артикул Google ученый
млн лет, Z., Guo, W., Guo, X., Wang, X., Kang, L. (2011) Модуляция фазовых изменений поведения мигрирующей саранчи путем метаболического пути катехоламинов. Proc. Natl. Акад. Sci. США 108 (10), 3882–3887
PubMed CAS PubMed Central Статья Google ученый
MacRae, T.H. (2010) Экспрессия генов, регуляция метаболизма и стрессоустойчивость во время диапаузы. Клетка. Мол. Life Sci. 67 , 2405–2424
PubMed CAS Статья Google ученый
Макинсон, Дж.К., Олдройд, Б.П., Шаерф, Т.М., Ваттаначайингчароен, В., Бикман, М. (2011) Переезд домой: выбор места для гнезда медоносной пчелы Красный карлик ( Apis florea ). Behav. Ecol. Sociobiol. 65 (5), 945–958
Артикул Google ученый
Мартин П. (1963) Die Steuerung der Volksteilung beim Schwärmen der Bienen. Zugleich ein Beitrag zum Problem der Wanderschwärme. Insectes Soc. 10 , 13–42
Артикул Google ученый
Мацука, М., Verma, L.R., Wongsiri, S., Shrestha, K.K., Partap, U (eds) (1998) Азиатские пчелы и пчеловодство: прогресс исследований и разработок. В: Материалы четвертой Международной конференции Азиатской ассоциации пчеловодства, Катманду, 23–28 марта 1998 г., Science Publishers, Inc., Энфилд, штат Нью-Хэмпшир. 274 p.
Mattila, H.R., Otis, G.W. (2007) Истощение ресурсов пыльцы вызывает переход к бесплодным популяциям долгоживущих пчел каждую осень. Ecol. Энтомол. 32 , 496–505
Артикул Google ученый
Маурицио А.(1950) Влияние кормления пыльцой и выращивания расплода на продолжительность жизни и физиологическое состояние медоносной пчелы: предварительный отчет. Пчелиный мир 31 , 9–12
Google ученый
McGlynn, T.P. (2012) Экология передвижения гнезд социальных насекомых. Анну. Преподобный Энтомол. 57 , 291–308
PubMed CAS Статья Google ученый
МакИндо, Н.Э. (1915) Орган медоносной пчелы, производящий запах. Proc. Акад. Natl. Sci. Phila. 66 , 542–555
Google ученый
Melathopoulous, A.P., Winston, M.L., Pettis, J.S., Pankiw, T. (1996) Влияние феромона нижней челюсти матки на инициацию и поддержание маточников у медоносной пчелы ( Apis mellifera L.). Может. Энтомол. 128 , 263–272
Артикул Google ученый
Михельсен, А., Кирхнер, У.Х., Андерсен, Б.Б., Линдауэр, М. (1986) Сигналы вибрации кряканья и кряканья пчелиных маток: количественный анализ. J. Comp. Physiol. A. 158 (5), 605–611
Статья Google ученый
Michener, C.D. (2013) Мелипонини. В: Vit, P., Pedro, S.R.M., Roubik, D.W. (ред.) Горшечный мед: наследие безжалостных пчел, стр. 3–18. Спрингер, Нью-Йорк
Google ученый
Moneti, G., Дани, Ф.Р., Пиераччини, Г., Туриллацци, С. (1997) Твердофазная микроэкстракция эпикутикулярных углеводородов насекомых для газохроматографического / масс-спектрометрического анализа. Rapid Commun. Mass Sp. 11 , 857–862
CAS Статья Google ученый
Морс, Р.А. (1963) Ориентация роя медоносных пчел. Наука 141 , 357–358
PubMed CAS Статья Google ученый
Науманн, К., Уинстон, М.Л., Слессор, К. (1993) Движение медоносной пчелы ( Apis mellifera L.) с нижнечелюстным феромоном королевы в густонаселенных и малонаселенных колониях. J. Поведение насекомых. 6 (2), 211–223
Артикул Google ученый
Nieh, J.C. (1993) Стоп-сигнал медоносных пчел: пересмотр своего сообщения. Behav. Ecol. Sociobiol. 33 , 51–56
Артикул Google ученый
Ние, Дж.C. (1998) Сигнал сотрясения медоносной пчелы: функция и конструкция модулирующего сигнала связи. Behav. Ecol. Sociobiol. 42 (1), 23–36
Артикул Google ученый
Отис, Г.В., Уинстон, М.Л., Тейлор, О.Р. (1981) Наполнение и рассредоточение африканизированных стаей медоносных пчел. J. Apic. Res. 20 (1), 3–11
Google ученый
Отт, С.Р., Верлинден, Х., Роджерс, С.М., Брайтон, К.Х., Куа, П.С., Влейгельс, Р.К., Вердонк, Р., Ванден Брок, Дж. (2012) Proc. Natl. Акад. Sci. США 109 (7), E381 – E387
PubMed CAS PubMed Central Статья Google ученый
Page, R.E., Peng, C.Y.S. (2001) Старение и развитие социальных насекомых с упором на медоносную пчелу, Apis mellifera L. Exp. Геронтол. 36 , 695–711
PubMed Статья Google ученый
Пейдж, Р.Э., Блюм, М.С., Фалес, Х.М. (1988) о-аминоаэтофенон, феромон, отпугивающий медоносных пчел ( Apis mellifera L.). Experientia 44 (3), 270–271
PubMed CAS Статья Google ученый
Пейдж-младший, Р.Э., Рюппелл, О., Амдам, Г.В. (2012) Генетика воспроизводства и регуляция социального поведения медоносной пчелы ( Apis mellifera L.). Анну. Преподобный Жене. 46 , 97–119
PubMed CAS Статья Google ученый
Художник-Курт, С., Шнайдер, С.С. (1998) Возраст и поведение медоносных пчел, Apis mellifera (Hymenoptera: Apidae), которые передают сигналы вибрации маткам и маточникам. Этология 104 , 475–485
Статья Google ученый
Петерс, К., Ито, Ф. (2001) Расселение колоний и эволюция морфологии матки у социальных перепончатокрылых. Анну. Преподобный Энтомол. 46 , 601–630
PubMed CAS Статья Google ученый
Петтис, Дж.С., Хиго, Х.А., Панкив, Т., Уинстон, М.Л. (1997) Королева, разводящая медоносную пчелу, — свидетельство сигнала о плодовитости. Insectes Soc. 44 , 311–322
Артикул Google ученый
Пикетт, Дж. А., Уильямс, И. Х., Мартин, А. П., Смит, М. К. (1980) Феромон Насонова медоносной пчелы, Apis mellifera L. (Hymenoptera: Apidae) Часть I. Химическая характеристика. J. Chem. Ecol. 6 , 425–434
CAS Статья Google ученый
Пирс, А.Л., Льюис, Л.А., Шнайдер, С.С. (2007) Использование сигнала вибрации и поведения рабочего трубопровода во время роения медоносных пчел, Apis mellifera . Этология 113 (3), 267–275
Статья Google ученый
Квеллер, округ Колумбия (2008) Поведенческая экология: социальная сторона диких дрожжей. Природа 456 , 589–590
PubMed CAS Статья Google ученый
Ранжел Дж., Сили, Т.Д. (2008) Сигналы, инициирующие массовый исход пчелиного роя из своего гнезда. Anim. Behav. 76 , 1943–1952
Артикул Google ученый
Рангел, Дж., Сили, Т. Д. (2012) Деление колоний у медоносных пчел: размер и значение фракции роя. Insectes Soc. 59 , 453–462
Артикул Google ученый
Ранжел Дж., Маттила, Х.Р., Сили, Т.Д. (2009) Отсутствие внутриколониального кумовства во время деления колонии у медоносных пчел. Proc. R. Soc. Лонд., Биол. Sci. 276 , 3895–3900
Артикул Google ученый
Ранжел, Дж., Гриффин, С.Р., Сили, Т.Д. (2010) Олигархия разведчиков гнездовых участков вызывает уход пчелиного роя из улья. Behav. Ecol. Sociobiol. 64 (6), 979–987
Артикул Google ученый
Ритчоф, К.К., Сили, Т.Д. (2008) Жужжание: как пчелы сигнализируют: «Пора идти! Anim. Behav. 75 , 189–197
Артикул Google ученый
Робинсон, Г.Э. (1992) Регулирование разделения труда в сообществах насекомых. Анну. Преподобный Энтомол. 37 , 637–665
PubMed CAS Статья Google ученый
Робинсон, Г.Э., Ферналд, Р.Д., Клейтон, Д.Ф. (2008) Гены и социальное поведение. Science 322 (5903), 896–900
PubMed CAS PubMed Central Статья Google ученый
Рубик Д.В. (2006) Биология гнездования пчел без жала. Apidologie 37 , 124–143
Статья Google ученый
Руттнер, Ф. (1987) Биогеография и таксономия медоносных пчел. Спрингер, Нью-Йорк
Google ученый
Шмидт, Дж.О. (2001) Иерархия аттрактантов для пчелиных роев. J. Поведение насекомых. 14 , 469–477
Артикул Google ученый
Шмидт, Дж. О., Слессор, К. Н., Уинстон, М. Л. (1993) Роль Насонова и феромонов матки в привлечении пчелиных стаей. Naturwissenschaften 80 , 573–575
CAS Статья Google ученый
Schmitt, U., Lubke, G., Francke, W. (1991) Секреция лапок отмечает источники пищи у шмелей (Hymenoptera: Apidae). Химиоэкология 2 (1), 35–40
CAS Статья Google ученый
Schneider, S.S. (1987) Модуляция активности рабочих вибрационным танцем медоносной пчелы, Apis mellifera . Этология 74 (3), 211–218
Статья Google ученый
Шнайдер, С.S. (1990a) Поведение королевы и взаимодействие рабочих маток в убегающих и роящихся колониях африканской медоносной пчелы, Apis mellifera scutellata (Hymenoptera, Apidae). J. Kansas Entomol. Soc. 63 (1), 179–186
Google ученый
Schneider, S.S. (1990b) Характеристики гнезд и поведение при пополнении покидающих колоний африканской медоносной пчелы Apis mellifera scutellata в Африке. Дж.Поведение насекомых. 3 (2), 225–240
Артикул Google ученый
Schneider, S.S. (1991) Модуляция активности матки с помощью танца вибрации в роящихся колониях африканской медоносной пчелы, Apis mellifera scutellata (Hymenoptera, Apidae). J. Kansas Entomol. Soc. 64 (3), 269–278
Google ученый
Шнайдер, С.С., ДеГранди-Хоффман, Г.(2008) Замена матки в колониях африканских и европейских пчел с послепогреванием и без него. Insectes Soc. 55 (1), 79–85
Артикул Google ученый
Шнайдер, С.С., Льюис, Л.А. (2004) Вибрационный сигнал, модуляционная коммуникация и организация труда у медоносных пчел, Apis mellifera . Apidologie 35 , 117–131
Статья Google ученый
Шнайдер, С.С., МакНалли, Л. (1991) Вибрационное танцевальное поведение рабочих медоносных пчел без матки, Apis mellifera (Hymenoptera: Apidae). J. Поведение насекомых. 4 (3), 319–332
Артикул Google ученый
Schneider, S.S., McNally, L.C. (1992) Факторы, влияющие на сезонные побеги в колониях африканской медоносной пчелы, Apis mellifera scutellata . Insectes Soc. 39 (4), 403–423
Артикул Google ученый
Шнайдер, С.С., МакНалли, Л. (1994) Танец покачивания, связанный с сезонным бегством в колониях африканской медоносной пчелы, Apis mellifera scutellata . Insectes Soc. 41 , 115–127
Артикул Google ученый
Schneider, S.S., Stamps, J.A., Gary, N.E. (1986) Вибрационный танец медоносной пчелы. I Связь, регулирующая добычу пищи в двух временных масштабах. Anim. Behav. 34 , 377–385
Артикул Google ученый
Шнайдер, С.С., Вишер, П.К., Камазин, С. (1998) Поведение танцоров виляния в роях медоносных пчел, Apis mellifera, (Hymenoptera: Apidae). Этология 104 , 963–972
Статья Google ученый
Шнайдер, С.С., Пейнтер-Курт, С., ДеГранди-Хоффман, Г. (2001) Роль вибрационного сигнала во время конкуренции маток в колониях медоносных пчел, Apis mellifera . Anim. Behav. 61 , 1173–1180
Артикул Google ученый
Шульц К.М., Пассино, К.М., Сили, Т.Д. (2008) Механизм управления полетом в роях медоносных пчел: тонкие направляющие или пчелы-полосатики? J. Exp. Bio. 211 , 3287–3295
Артикул Google ученый
Сили, Т. Д. (1979) Распространение вещества королевы курьерами в пчелиных семьях. Behav. Ecol. Sociobiol. 5 , 391–415
Артикул Google ученый
Сили Т.Д. (1982) Адаптивное значение графика возрастного полиэтиизма в пчелиных семьях. Behav. Ecol. Sociobiol. 11 , 287–293
Артикул Google ученый
Сили, Т. Д. (1995) Мудрость улья: социальная физиология семей медоносных пчел. Издательство Гарвардского университета, Кембридж
Сили, Т. Д. (2010) Пчелиная демократия. Princeton University Press, Princeton
Seeley, T.Д., Бурман С.С. (1999) Групповое принятие решений в стаях медоносных пчел. Behav. Ecol. Sociobiol. 45 , 19–31
Артикул Google ученый
Сили, Т.Д., Фелл, Р.Д. (1981) Производство вещества королевы в колониях медоносных пчел ( Apis mellifera ), готовящихся к рою (Hymenoptera: Apidae). J. Kansas Entomol. Soc. 54 (1), 192–196
Google ученый
Сили Т.Д., Тауц, Дж. (2001) Трубопровод рабочих в стае медоносных пчел и его роль в подготовке к старту. J. Comp. Physiol. A. 187 (8), 667–676
PubMed CAS Статья Google ученый
Сили, Т.Д., Висшер, П.К. (2004) Групповое принятие решений при выборе места для гнезд медоносными пчелами. Apidologie 35 (2), 101–116
Статья Google ученый
Сили Т.Д., Морс, Р.А., Вишер, П.К. (1979) Естественная история полета стаи медоносных пчел. Psyche 86 (2–3), 103–114
Статья Google ученый
Сили, Т.Д., Вайденмюллер, А., Кюнхольц, С. (1998) Сигнал сотрясения медоносной пчелы информирует рабочих о необходимости подготовиться к большей активности. Этология 104 , 10–26
Статья Google ученый
Сили Т.Д., Кляйнхенц, М., Буйок, Б., Тауц, Дж. (2003) Тщательная разминка перед взлетом в роях медоносных пчел. Naturwissenschaften 90 (6), 256–260
PubMed CAS Статья Google ученый
Сили, Т.Д., Вишер, П.К., Пассино, К.М. (2006) Групповое принятие решений в пчелиных роях. Являюсь. Sci. 94 (3), 220–229
Артикул Google ученый
Сили Т.D., Visscher, P.K., Schlegel, T., Hogan, P.M., Franks, N.R., Marshall, J.A.R. (2012) Стоп-сигналы обеспечивают перекрестное торможение коллективного принятия решений роем медоносных пчел. Science 335 (6064), 108–111
PubMed CAS Статья Google ученый
Sihag, R.C. (1998) Экобиология маленькой медоносной пчелы ( Apis florea ) в полузасушливом субтропическом климате Индии. В: М. Мацука, Л. Р. Верма, С. Вонгсири, К.К. Шреста и У. Партап (ред.) Азиатские пчелы и пчеловодство: прогресс исследований и разработок. Science Publishers, Enfield, NY, стр. 50–52
Simpson, J. (1957a) Наблюдения за колониями медоносных пчел, подвергнутых обработке, направленной на то, чтобы вызвать роение. Proc. R. Entomol. Soc. Лондон А. 32 (10–12), 185–192
Google ученый
Simpson, J. (1957b) Частота роения среди колоний медоносных пчел в Англии.J. Agr. Sci. 49 , 387–393
Артикул Google ученый
Simpson, J. (1958) Факторы, которые вызывают скопление колоний Apis mellifera . Insectes Soc. 5 (1), 77–95
Артикул Google ученый
Simpson, J. (1959) Изменения в частоте роения среди колоний Apis mellifera в течение всего лета.Insectes Soc. 6 (1), 85–99
Артикул Google ученый
Симпсон, Дж. (1963) Восприятие королевы пчелиными роями. Природа 199 , 94–95
Артикул Google ученый
Simpson, J., Riedel, I.B.M. (1964) Появление стай из колоний Apis mellifera . Поведение 23 , 140–148
Статья Google ученый
Слессор, К.Н., Камински, Л.-А., Кинг, Г.С.С., Борден, Дж., Уинстон, М. (1988) Семиохимические основы реакции свиты на пчелиных маток. Природа 332 , 354–356
CAS Статья Google ученый
Slessor, K.N., Winston, M.L., Le Conte, Y. (2005) Связь феромонов у медоносной пчелы ( Apis mellifera L.). J. Chem. Ecol. 31 (11), 2731–2745
PubMed CAS Статья Google ученый
Смит, К.Р., Тот, А.Л., Суарес, А.В., Робинсон, Г.Е. (2008) Генетический и геномный анализ разделения труда в сообществах насекомых. Nat. Преподобный Жене. 9 (10), 735–748
PubMed CAS Статья Google ученый
Салливан, Дж. П., Джассим, О., Фарбак, С. Е., Робинсон, Г. (2000) Ювенильный гормон влияет на развитие поведения взрослых медоносных пчел. Horm. Behav. 37 , 1–14
PubMed CAS Статья Google ученый
Самптер, Д.J.T. (2006) Принципы коллективного поведения животных. Фил. Пер. R. Soc. B 361 (1465), 5–22
PubMed CAS PubMed Central Статья Google ученый
Tarpy, D.R., Fletcher, D.J.C. (2003) «Опрыскивание» поведения во время соревнований маток медоносных пчел. J. Поведение насекомых. 16 , 425–437
Артикул Google ученый
Тот, А.Л., Робинсон, Г. (2005) Питание рабочих и разделение труда у медоносных пчел. Anim. Behav. 69 , 427–435
Артикул Google ученый
Тот, А.Л., Робинсон, Г.Э. (2007) Эво-дево и эволюция социального поведения. Тенденции Genet. 23 (7), 334–341
PubMed CAS Статья Google ученый
Velicer, G.J., Yu, Y.T.(2003) Эволюция нового кооперативного роения бактерии Myxococcus xanthus . Nature 425 (6953), 75–78
PubMed CAS Статья Google ученый
Венкатеш, Г., Редди, К.С. (1989) Скорость роения и побега гигантской медоносной пчелы, Apis dorsata F. Proc. Индийский акад. Sci. (Anim. Sci.) 98 (6), 425–430
Статья Google ученый
Виллелла, А., Холл, Дж. К. (2008) Нейрогенетика ухаживания и спаривания у Drosophila . Adv. Genet. 62 , 67–187
PubMed CAS Статья Google ученый
Висшер, П.К., Сили, Т.Д. (2007) Координация отъезда группы: кто подает сигналы по трубопроводу на стаи медоносных пчел? Behav. Ecol. Sociobiol. 61 , 1615–1621
Артикул Google ученый
Visscher, P.К., Шепардсон, Дж., Маккарт, Л., Камазин, С. (1999) Вибрационный сигнал модулирует поведение медоносных пчел при домашней охоте ( Apis mellifera ). Этология 105 , 759–769
Статья Google ученый
Вест-Эберхард, М.Дж. (1982) Природа и эволюция роения тропических социальных ос (Vespidae, Polistinae, Polybiini). В: П. Джейссон (ред.) Социальные насекомые в тропиках, т. I. С. 97–128. Univ. Paris XIII Press, Париж
Whitfield, C.В., Бехура, С. К., Берлохер, Ш., Кларк, А. Г., Джонстон, Дж. С., Шеппард, В. С., Смит, Д. Р., Суарес, А. В., Уивер, Д., Цуцуи, Н. Д. (2006a) Трижды из Африки: древние и недавние разложения медоносной пчелы, Apis mellifera . Наука 314 , 642–645
PubMed CAS Статья Google ученый
Whitfield, C.W., Ben-Shahar, Y., Brillet, C., Leoncini, I., Crauser, D., Le Conte, Y., Rodriguez-Zas, S., Робинсон, Г. (2006b) Геномный анализ поведенческого созревания медоносной пчелы. Proc. Natl. Акад. Sci. США 103 (44), 16068–16075
PubMed CAS PubMed Central Статья Google ученый
Wilson, E.O. (1971) Общества насекомых. Издательство Гарвардского университета, Кембридж
Google ученый
Winston, M.L. (1979) Демография внутри колонии и скорость воспроизводства африканизированных пчел в Южной Америке.Behav. Ecol. Sociobiol. 4 , 279–292
Артикул Google ученый
Winston, M.L. (1980) Рой, последующее потепление и воспроизводимость неуправляемых семей пчел ( Apis mellifera ). Insectes Soc. 27 (4), 391–398
Артикул Google ученый
Winston, M.L. (1987) Биология медоносной пчелы. Издательство Гарвардского университета, Кембридж
Google ученый
Уинстон М.Л., Тейлор О. (1980) Факторы, предшествующие выращиванию матки африканизированной пчелы ( Apis mellifera ) в Южной Америке. Insectes Soc. 27 (4), 289–304
Артикул Google ученый
Winston, M.L., Dropkin, J.A., Taylor, O.R. (1981) Демография и характеристики жизненного цикла двух рас медоносных пчел ( Apis mellifera ). Oecologia 48 (3), 407–413
Статья Google ученый
Уинстон М.Л., Слессор, К.Н., Смирле, М.Дж., Кандил, А.А. (1982) Влияние производимого маткой вещества, 9HDA, на поведение роя у медоносной пчелы Apis mellifera L. J. Chem. Ecol. 8 (10), 1283–1288
PubMed CAS Статья Google ученый
Winston, ML, Slessor, KN, Willis, LG, Naumann, K., Higo, HA, Wyborn, MH, Kaminski, LA (1989) Влияние феромонов нижней челюсти королевы на привлечение рабочих к скоплениям роя и ингибирование разведение матки в медоносной пчеле ( Apis mellifera L.). Insectes Soc. 36 (1), 15–27
Артикул Google ученый
Winston, M.L., Higo, H.A., Colley, S.J., Pankiw, T., Slessor, K.N. (1991) Роль нижнечелюстного феромона королевы и скопление колоний в репродуктивном роении медоносных пчел ( Apis mellifera L.) (Hymenoptera: Apidae). J. Поведение насекомых. 4 (5), 649–660
Артикул Google ученый
Восслер, Т.К., Крю Р. (1999a) Секреция тергальных желез королевы медоносных пчел влияет на развитие яичников у рабочих, содержащихся в клетке. Apidologie 30 , 311–320
Статья Google ученый
Wossler, T.C., Crewe, R.M. (1999b) Выделяющие эффекты секреции тергальных желез маток пчел ( Apis mellifera L.). J. Поведение насекомых. 12 , 343–350
Артикул Google ученый
Цзэн, З., Huang, Z.H., Qin, Y., Pang, H. (2005) Титры ювенильного гормона гемолимфы у рабочих медоносных пчел в нормальных и предварительных условиях. J. Econ. Энтомол. 98 (2), 274–278
PubMed CAS Статья Google ученый
Сон нарушает высокоуровневый синтаксический анализ речи, несмотря на значительную базовую обработку слуха
Главный новый результат, о котором здесь сообщается, заключается в том, что во время сна базовое нейронное кодирование акустических характеристик речи сохраняется, тогда как синтаксический анализ лингвистических структур более высокого порядка очевиден для внятная речь при бодрствовании, нарушена.Эти результаты расширяют наше понимание возможностей и ограничений корковой обработки во время сна и демонстрируют эффективность подхода CHT для зондирования когнитивных процессов высокого уровня, скрытых в неотзывчивых состояниях.
Изучение лингвистической обработки во время бодрствования с помощью CHT
Наши результаты по бодрствованию согласуются с данными Ding et al. (2015), показав, что корковая активность одновременно отслеживает динамику языковых структур на нескольких иерархических уровнях, когда речь разборчива.При неразборчивой речи корковые реакции проявляются только на акустической / слоговой скорости. Наши результаты расширяют исходные данные по нескольким направлениям. Во-первых, использование двухсложных слов позволило различать слоговые (4 Гц) и словесные (2 Гц), что позволило разделить акустический и лингвистический аспекты речи. Во-вторых, наш дизайн не включал явную задачу, демонстрирующую, что пассивного слушания достаточно для выявления иерархического нейронного отслеживания языковых структур.В-третьих, наши результаты демонстрируют, что можно эффективно количественно оценить иерархический лингвистический синтаксический анализ с помощью неинвазивного, недорогого, портативного и легкодоступного ЭЭГ. Эти расширения открывают путь к использованию подхода CHT в широком спектре настроек, начиная от овладения языком и заканчивая исследованиями остаточной когнитивной обработки в различных клинических группах.
Глубина обработки речи во время сна
Текущие результаты вносят существенный вклад в наше понимание степени обработки речи во время сна.Мы обнаружили, что нейронная реакция на слоговой скорости, которая, вероятно, отражает основное слуховое представление акустической оболочки, была сопоставима во время бодрствования, быстрого сна и медленного сна. Это согласуется с предыдущими исследованиями, подтверждающими сохраненные ответы низкоуровневой слуховой коры во время сна (Issa and Wang, 2008; Nir et al., 2015). В то же время во время сна нарушается анализ лингвистических структур более высокого порядка, что подразумевает наличие функционального «узкого места», препятствующего полной обработке непрерывной речи.
Прежде чем обсуждать более широкие последствия, мы обращаемся к важной методологической оговорке: приводит ли отсутствие наблюдаемых низкочастотных пиков во время сна из-за повышенного фонового шума к плохой чувствительности, а не к недостатку высокоуровневого синтаксического анализа речи? Эта потенциальная критика относится в основном к медленному сну, потому что спонтанная активность во время быстрого сна похожа на бодрствование. Хотя применение трансформации CSD эффективно снижало медленную широко распространенную активность в NREM-сне (Fig. 3 D ), она, тем не менее, была выше, чем в бодрствовании.Однако мы полагаем, что это не объясняет наши результаты по двум причинам. Во-первых, наш экспериментальный план намеренно сосредоточен на тестировании различных речевых условий в каждом состоянии, в которых совпадают SNR и другие свойства сигнала. Во-вторых, мы провели несколько анализов с учетом различий SNR в разных штатах. Они показали, что, хотя акустические отклики с частотой 4 Гц существенно не различались между состояниями, ответы на уровне слова и предложения были обнаружены для разборчивой речи только во время бодрствования, на что указывают значимые взаимодействия между состоянием × частота.Тем не менее, мы признаем, что, учитывая преобладание низкочастотной фоновой активности во время медленного сна (как и в других невосприимчивых состояниях, таких как анестезия и вегетативные состояния), результаты следует интерпретировать с большой осторожностью и применять соответствующие меры контроля в будущих исследованиях.
Какова природа функционального узкого места, наблюдаемого во время сна между базовой обработкой слуха и лингвистическим анализом высокого уровня? Предыдущие результаты ЭКоГ с использованием парадигмы CHT приписывают фразовые и предложения фразовые ответы бессенсорным областям, участвующим в языковой обработке; например, левая IFG и двусторонняя TPJ области.Напротив, ответы на скорость слога были обнаружены в основном в слуховой коре (Ding et al., 2016). Отсутствие доказательств нейронного отслеживания речевых структур высокого уровня во время сна предполагает, что это нарушает эффективное распространение сигнала от слуховой коры к более высоким кортикальным областям. Таким образом, межкортикальные связи во время сна ограничиваются кратковременным электромагнитным возмущением (Massimini et al., 2005, 2007; Pigorini et al., 2015). Точно так же несколько исследований фМРТ продемонстрировали устойчивое ослабление речевых ответов в IFG и лобных областях во время сна (Portas et al., 2000; Dehaene-Lambertz et al., 2002; Wilf et al., 2016). Вместе с текущими результатами эти исследования предполагают, что обработка речи во время сна ограничивается акустической обработкой низкого уровня с сопоставимыми нейронными реакциями для разборчивой и неразборчивой речи.
Однако другие исследования сообщают о доказательствах остаточной семантической обработки во время сна при предъявлении отдельных слов (Kouider et al., 2014; Andrillon et al., 2016), пар слов (Brualla et al., 1998) и коротких предложений ( Ibáñez et al., 2006; Daltrozzo et al., 2012), хотя такая обработка обычно слабее и с измененной временной динамикой по сравнению с бодрствованием (Brualla et al., 1998; Perrin et al., 2002). Пытаясь согласовать эти, казалось бы, противоречивые результаты, мы предполагаем, что функциональное узкое место для обработки речи во время сна — это не семантический анализ как таковой, а скорее опосредующий процесс сегментации. Одним из ключевых отличий этого исследования от предыдущих является использование непрерывной речи, а не отдельных слов или коротких предложений.Это не только позволило нам преодолеть серьезные методологические ограничения, такие как распространенность К-комплексов, но и задействовать процесс сегментации слов, который является предпосылкой для непрерывного понимания речи. Действительно, обработка речи критически зависит от точного разбора текущего входящего акустического потока на дискретные и значимые языковые единицы (Hickok et al., 1993; Mattys, 1997; Giraud and Poeppel, 2012; Doelling et al., 2014). Сам по себе синтаксический анализ представляет собой сложный иерархический когнитивный процесс (Greenberg et al., 2004; Ghitza, 2012), который включает сопоставление восходящих сигналов с существующими лексическими, синтаксическими и семантическими представлениями (Traxler, 2014), а также процессы прогнозирования и упреждения (Arnal et al., 2011; DeLong et al., 2014). Текущие результаты показывают, что, как минимум, во время сна нарушается процесс сегментации слов, процесс, который требует постоянного анализа непрерывного глобального потока ввода (Strauss et al., 2015; Tononi et al., 2016) . Таким образом, остается возможным, что, когда фразы или предложения представлены в дискретном формате (как в большинстве предыдущих исследований), отдельные слова могут быть идентифицированы и полностью обработаны до семантического уровня даже во время сна.
Еще одним важным аспектом синтаксического анализа непрерывной речи является интеграция информации в нескольких временных масштабах (Rosen, 1992; Greenberg et al., 2003; Ghitza, 2012; Zion Golumbic et al., 2013), что позволяет получать информацию в коротком масштабе (фонемы / слоги ) для создания лексических единиц (например, слов) и структур более высокого порядка (словосочетаний и предложений). Такая интеграция требует буферов краткосрочной памяти для хранения информации в течение более длительных периодов; например, чтобы справиться с большими «временными окнами восприятия» (Lerner et al., 2011; Луо и Поппель, 2012; Chait et al., 2015). Примечательно, что предыдущие исследования показали, что нервная активность во время сна ограничена короткими временными рамками в диапазоне сотен миллисекунд, что предотвращает возникновение длительных причинно-следственных связей (Pigorini et al., 2015; Strauss et al., 2015). Предлагаемое ограничение мозговой активности во время сна короткими интервалами времени также может способствовать наблюдаемому здесь уменьшенному иерархическому синтаксическому анализу непрерывной речи.
Одним из важных ограничений текущего исследования является то, что просодические сигналы были намеренно удалены из речевого материала, что позволило нам исследовать грамматический синтаксический анализ речи независимо от коррелированных акустических вариаций.Однако, поскольку синтаксический анализ естественной речи также выигрывает от просодии (Eckstein and Friederici, 2006), еще предстоит проверить, может ли просодия облегчить непрерывный анализ речи во время сна.
Сон и невнимательность
Результаты, аналогичные тем, которые были обнаружены здесь во время сна, недавно были получены для речи без участия оператора, для которой нейронное отслеживание устойчиво в слуховой коре, но существенно снижается в областях языка высшего порядка (Zion Golumbic et al., 2013 ; Rimmele et al., 2015). Таким образом, акустические особенности речи без участия оператора влияют на поведение в большей степени, чем высокоуровневые (семантические) характеристики (Ellermeier et al., 2015; Wöstmann and Obleser, 2016). Дополнительные сходства между сном и невниманием существуют на поведенческом уровне, где речь, оставленная без внимания, обычно не может быть открыто вспомнить (Lachter et al., 2004), хотя некоторые слова, относящиеся к личному, иногда привлекают внимание (Cherry, 1953; Moray, 1958, 1959; Bentin et al., 2004). al., 1995; Wood and Cowan, 1995), как и во время сна.Эти сходства между обработкой речи во время сна и невниманием могут иметь более широкое значение в отношении природы функциональных «узких мест» в обработке речи (Broadbent, 1958; Treisman, 1969), составляя интригующую тему для будущих исследований.
Выводы
Насколько нам известно, это исследование является первым исследованием иерархического анализа непрерывной речи во время сна. Мы обнаружили, что обработка слуха снизу вверх сохраняется во сне и сравнима с таковой в бодрствовании.В отличие от этого, нейронное отслеживание лингвистических структур высшего порядка — слов, фраз и предложений — нарушается во сне подобно тому, как это происходит при разговоре без внимания или неразборчивой речи во время бодрствования. Текущие результаты показывают, что анализ непрерывной речи, который требует интеграции в нескольких временных масштабах, сопоставления восходящего ввода с сохраненными лингвистическими представлениями и нисходящего прогнозирующего кодирования, может быть невозможен без открытого внимания и осознанности. Наши результаты предполагают функциональный барьер между слуховыми ощущениями и лингвистической обработкой, барьер, который может иметь важное значение для обеспечения сохранения сна перед лицом внешних событий и для поддержки его функций.Это исследование закладывает основу для изучения остаточной обработки речи при различных состояниях сознания, анестезии, нейродегенерации, развитии и языковых расстройствах.
PLOS ONE: Аудиосистема
Хуэй Ван, Бэй Ли, […], Шанкай Инь
Роберт Вундерлих, Пиа Лау, […], Христо Пантев
Насим А.Чоудхури, Джессика А. Параскандо, Апрель А. Бенасич
Рик Л. Дженисон, Ричард А. Реале, […], Мэтью А. Ховард III
Тэцуя Сига, Хайке Альтен, […], Карлес Эскера
Такаши Уэяма, Томохиро Дониси, […], Йошики Канеоке
Шрирам Бутхалингам, Крис Аллан, Пруденс Аллен, Дэвид Перселл
Юэсинь Цай, Ицин Чжэн, […], Гуйшэн Чен
Эмма Лейшман, Брайан Ф. О’Доннелл, […], Сандра Л. Морзорати
Хосе А. Гарсиа-Лазаро, Кэтрин Н. Шепард, […], Николай А. Лесика
Стэнли Шефт, Валерий Шафиро, […], Радж С. Шах
Блейк Э.Дворецкий, Эйми Дж. Холл, Стивен Дж. Ломбер
% PDF-1.4 % 624 0 объект > эндобдж xref 624 86 0000000015 00000 н. 0000002121 00000 н. 0000002305 00000 н. 0000003029 00000 н. 0000003732 00000 н. 0000003874 00000 н. 0000004416 00000 н. 0000005021 00000 н. 0000005472 00000 н. 0000009976 00000 н. 0000010234 00000 п. 0000011714 00000 п. 0000011826 00000 п. 0000012347 00000 п. 0000012592 00000 п. 0000012652 00000 п. 0000012814 00000 п. 0000013092 00000 п. 0000013333 00000 п. 0000037687 00000 п. 0000038616 00000 п. 0000038877 00000 п. 0000038926 00000 п. 0000052398 00000 п. 0000052934 00000 п. 0000052987 00000 п. 0000053261 00000 п. 0000053589 00000 п. 0000053861 00000 п. 0000053959 00000 п. 0000054267 00000 п. 0000056262 00000 п. 0000056425 00000 п. 0000056869 00000 п. 0000057221 00000 п. 0000057375 00000 п. 0000057416 00000 п. 0000057482 00000 п. 0000057638 00000 п. 0000057679 00000 п. 0000069131 00000 п. 0000089152 00000 п. 0000089277 00000 н. 0000089809 00000 п. 0000089970 00000 н. 00000
00000 п. 00000 00000 п. 00000
00000 н. 00000 00000 п. 00000
00000 п. 00000 00000 п.